
Chytridiomycota
| Chytridiomycota Chytridiomycotina | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Classificação científica | |||||||||
| |||||||||
| Classes | |||||||||




Chytridiomycota (grego χυτρίδιον; khutrídion; "pequeno pote", alusão à estrutura que contém os zoósporos) é uma divisão do reino Fungi que inclui um largo grupo de organismos zoospóricos heterotróficos, essencialmente aquáticos, informalmente conhecidos por fungos quitrídios, caracterizados por apresentarem paredes celulares quitinosas e apresentarem nutrição por absorção. Os quitrídios são uma das primeiras linhagens de fungos que divergiram, com a participação no reino Fungi demonstrada pela presença de quitina na parede celular, flagelos com chicote posterior, nutrição por absorção, uso de glicogénio como composto de armazenamento de energia e síntese de lisina pela via α-amino-ácido-adípico (AAA).[4][5] Neste agrupamento taxonómico todas as espécies possuem um estágio zoospórico no seu ciclo de vida,[6] sendo o único clado de verdadeiros fungos (Eumycota) que se reproduzem com recurso à formação de zoósporos.[7] A divisão é um táxon monotípico tendo Chytridiomycotina como única subdivisão, repartida por 9 classes.[8] Os quitrídios são sapróbicos, capazes de degradar materiais refratários como quitina e queratina, embora por vezes possam agir como parasitas.[9] Houve um aumento significativo na investigação sobre os quitrídios desde a descoberta de Batrachochytrium dendrobatidis, o agente causal da quitridiomicose.[10][11]
Descrição[editar | editar código-fonte]
A divisão Chytridiomycota tem vindo a sofrer um conjunto de profundas revisões taxonómicas, a última das quais em 2018,[1] da qual resultou o reconhecimento de uma única subdivisão, Chytridiomycotina, nela se incluindo as 9 classes agora reconhecidas com base em estudos de filogenética molecular. Alguns géneros continuam em incertae sedis, o que seguramente levará novas revisões da taxonomia do grupo. Na sus presente circunscrição taxonómica, os Chytridiomycota constituem um clado que agrupa a maioria dos membros do reino Fungi que produzem células móveis (os restantes estão nas divisões Blastocladiomycota e Aphelidiomycota).[12]
Com exceção das poucas espécies que produzem células poliflageladas, os zoósporos e gâmetas destes organismos são células móveis que apresentam um único flagelo eucariótico do tipo chicote, inserido na sua parte posterior, sendo esta a sua principal característica morfológica distintiva.[13]
Os fungos pertencentes a este agrupamento são similares aos fungos superiores na composição de suas paredes celulares e pela bioquímica celular, semelhança que é confirmada pela análise sequencial de um conjunto de subunidades ribossómicas. Mesmo sem se considerar aspectos bioquímicos ou moleculares, há espécies filamentosas entre os Chytridiomycota que demonstram a relação com os fungos superiores.[14]. Outras características que possuem em comum com os restantes fungos do clado Eumycota são: a estrutura cenocítica do talo e a conversão do zigoto em esporo ou esporângio de resistência, ou no desenvolvimento de talo diploide. Possuem talo de desenvolvimento endógeno ou exógeno, holocárpico ou eucárpico, monocêntrico, policêntrico, rizoidal ou micelial; paredes celulares com quitina e glucanos; cristas mitocondriais achatadas; divisões intranucleares cêntricas; zoósporos sem mastigonemas ou escamas (flagelo do tipo chicote), com um único sistema de raízes flagelares e algumas vezes rumpossomas. A reprodução assexual ocorre por meio de zoósporos e a reprodução sexual por copulação planogamética, copulação de gametângios ou somatogamia.[15][13]
Morfologia e reprodução[editar | editar código-fonte]
A reprodução sexuada é comum e bem conhecida entre os membros dos quitrídios, muitos dos quais praticam uma versão de oogamia em que o macho é móvel e a fêmea é estacionária. Esta é a primeira ocorrência de oogamia que se conhece no reino Fungi.[5] Uma vez fertilizado, o zigoto origina um oósporo encistado ou móvel,[9] que finalmente se torna um esporo de resistência que posteriormente germina e dá origem a novos zoosporângios.[5]
Após a liberação do esporo de resistência germinado, os zoósporos procuram um substrato adequado para crescimento usando quimiotaxia ou fototaxia. Algumas espécies enquistam e germinam diretamente sobre o substrato; outras enquistam e germinam a uma curta distância. Uma vez germinado, as enzimas libertadas do zoósporo começam a decompor o substrato, utilizando os materiais e a energia para produzir um novo talo. Os talos são cenocíticos e geralmente não formam micélio verdadeiro (tendo rizoides em substituição).
Os quitrídios apresentam vários padrões de crescimento diferentes. Alguns são holocárpicos, o que significa que eles só produzem zoosporângios e zoósporos. Outros são eucárpicos, ou seja, produzem outras estruturas, como rizoides, além do zoosporângio e zoósporos. Alguns quitrídios são monocêntricos, o que significa que um único zoósporo dá origem a um único zoosporângio. Outros são policêntricos, o que significa que um zoósporo dá origem a vários zoosporângios conectados por um rizomicélio. Os rizoides não têm núcleos, enquanto um rizomicélio os pode ter.[5]
O crescimento continua até que um novo grupo de zoósporos esteja pronto para ser libertado. Os quitrídios têm um conjunto diversificado de mecanismos de libertação de esporos que podem ser agrupados nas amplas categorias de operculados ou inoperculados. A descarga operculada envolve o desprendimento completo ou incompleto de uma estrutura semelhante a uma tampa, chamada opérculo, permitindo que os zoósporos saiam do esporângio. Os quitrídios inoperculados libertam os zoósporos através de poros, fendas ou papilas.[9]
Ciclo de vida[editar | editar código-fonte]
Os quitrídios são variadas em sua forma de vida, em suas interações sexuais e principalmente no ciclo de vida. A meiose e a mitose lembram os mesmos processos ocorrentes em outros fungos. São intranucleares, ou seja, o envoltório permanece intacto até à telófase tardia. Distinguem-se dos outros fungos principalmente pelas células móveis características (zoósporos e gâmetas), a maioria contendo apenas um único flagelo liso e posterior. Os quitrídios exibem vários modos de reprodução, algumas não desenvolvem micélio e outras tem rizoides finos.
A reprodução assexuada ocorre por zoósporos produzidos em esporângios e os esporângios podem ter opérculos ou ser inoperculados. A reprodução sexuada é conhecida em poucas espécies, sendo que a mesma pode ser por copulação gametangial, copulação planogamética (iso, aniso ou oogâmica) ou somatogamia.[16]
-
 Tipos de talos de quitrídios.
Tipos de talos de quitrídios. -
 Descarga de zoósporos do género Phylctochytrium sp.
Descarga de zoósporos do género Phylctochytrium sp. -
 Esporângio e zoósporos dos quitrídios da espécie B. dendrobatidis, vista microscópio eletrónico de varredura.
Esporângio e zoósporos dos quitrídios da espécie B. dendrobatidis, vista microscópio eletrónico de varredura. -
Esporângios e zoósporos de quitrídios vistos ao microscópio.
.png)
.jpg)

Os Chytridiomycota são incomuns entre os Fungos, pois produzem zoósporos como principal forma de reprodução.[9][17] Para a maioria dos membros de Chytridiomycota, a reprodução sexuada não é conhecida. A reprodução assexuada ocorre através da libertação de zoósporos (presumivelmente) derivados da mitose.[9]
Onde foi descrita, a reprodução sexuada dos quitrídios ocorre através de uma variedade de métodos. Naquele que é o ciclo mais comum, o zigoto resultante forma um esporo de resistência, que funciona como um meio de sobreviver a condições adversas.[9] Em alguns membros, a reprodução sexual é conseguida através da fusão de isogâmetas (gâmetas do mesmo tamanho e forma). Este grupo inclui os notáveis patógenos de plantas pertencentes ao género Synchytrium. Alguns parasitas de algas praticam oogamia, processo no qual um gâmeta masculino móvel liga-se a uma estrutura imóvel contendo o gâmeta feminino. Em outro grupo, dois talos produzem tubos que se fundem e permitem que os gâmetas se encontrem e se fundam.[9] Neste último grupo, rizoides de estirpes compatíveis são mutuamente atraídos e na fusão ambos os núcleos migram para fora do zoosporângio e para os rizoides unidos, onde também se fundem. O zigoto resultante germina num esporo de resistência.[4]
-
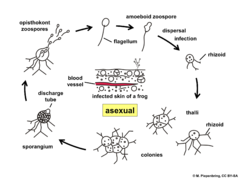 Ciclo de vida de Batrachochytrium dendrobatidis.
Ciclo de vida de Batrachochytrium dendrobatidis. -
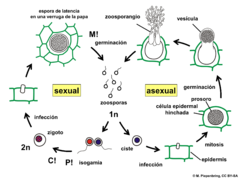 Ciclo de vida de Synchytrium endobioticum na batateira.
Ciclo de vida de Synchytrium endobioticum na batateira. -
 Synchytrium endobioticum sobre batatas.
Synchytrium endobioticum sobre batatas.
.png)
.png)

Distribuição e ecologia[editar | editar código-fonte]
Os fungos pertencentes a este agrupamento têm distribuição natural do tipo cosmopolita, presentes nos ecossistemas terrestres e aquáticos, a maioria como sapróbios, embora um número alargado seja parasita de algas, anfíbios, outros fungos, animais microscópicos e plantas superiores.[6]
Os membros do grupo ocorrem em habitats diversificados, desde os trópicos às regiões árticas.[18] Ocorrem em sistemas aquáticos tais como córregos, lagoas e zonas estuarinas (ambiente aquático de transição entre um rio e o mar), estando presentes ecossistemas marinhos, principalmente como parasitas de algas e de outros organismos componentes do plâncton. A maioria das espécies parece ocorrer em habitats terrestres, tais como florestas, solos agrícolas e mesmo em ambientes desérticos e turfeiras ácidas, onde a maioria vive como saprobiontes de substratos refratários à decomposição.[19]
Esse pendor para a saprobiose faz com que, provavelmente, a função ecológica mais importante desempenhada pelos quitrídios seja a decomposição.[14] São organismos omnipresentes e cosmopolitas, responsáveis na maioria dos ecossistemas pela decomposição de materiais refratários, como pólen, celulose, quitina e queratina.[14][9]
A capacidade de decompor materiais resistentes ao ataque biológico levou ao aparecimento de adaptações específicas, como a existência de quitrídios que vivem e crescem no pólen, ligando-se aos aos grãos de pólen por estruturas semelhantes a fios, chamadas rizoides.[20] Isso ocorre principalmente durante a reprodução assexuada, quando os zoósporos que se ligam ao pólen se reproduzem continuamente e formam novos quitrídios que se ligam a outros grãos de pólen para obter nutrientes. Essa colonização do pólen ocorre durante a primavera, quando o pólen que cai das árvores e plantas se acumula na superfície dos corpos de água, habitat preferido destes fungos.[9]
Habitats[editar | editar código-fonte]
Os quitrídios são fungos aquáticos, embora aqueles que prosperam na rede capilar em torno das partículas do solo sejam tipicamente considerados terrestres.[14][9] O zoósporo é principalmente um meio de explorar minuciosamente um pequeno volume de água para encontrar um substrato adequado, em vez de um meio de dispersão de longo alcance.[21]
Os quitrídios foram isolados de uma variedade de habitats aquáticos, incluindo turfeiras, pântanos, rios, lagoas, nascentes e fossos, mas também em habitats terrestres, como solos ácidos, solos alcalinos, solos de florestas temperadas, solos de florestas tropicais, solos árticos e antárticos.[14][9] Isso levou à crença de que muitas espécies de quitrídios são ubíquos e cosmopolitas.[14][9] No entanto, trabalhos taxonómicos recentes demonstraram que essas morfoespécies omnipresente e cosmopolitas escondem uma diversidade críptica nos níveis genético e ultraestrutural.[22][23]
Inicialmente, acreditou-se que os quitrídios aquáticos (e outros fungos zoospóricos) eram ativos principalmente no outono, inverno e primavera.[9] No entanto, inventários moleculares recentes de lagos recolhidos durante o verão indicam que os quitrídios são uma parte ativa e com elevada diversidade da comunidade microbiana eucariótica.[24]
Um dos ambientes terrestres menos esperados em que os quitrídios prosperassem são os solos periglaciais.[25] Comprovou-se que a população das espécies de Chytridiomycota pode ser mantida, embora haja falta de vida vegetal nessas regiões congeladas, devido à grande quantidade de água no solo periglacial e ao pólen arrastado pelos ventos desde as florestas situadas abaixo da linha de árvores (a timberline).
Parasitismo[editar | editar código-fonte]
Os quitrídios infetam principalmente algas e outros micróbios eucarióticos e procarióticos. Em meios lênticos, especialmente em pequenas lagoa de baixa profundidade, a infeção pode ser tão severa a ponto de controlar a produção primária.[5][26] Em consequência, tem sido sugerido que os quitrídios parasitas têm um grande efeito sobre as cadeias alimentares de lagos e lagoas.[27]
Os quitrídios também podem infetar espécies de plantas, tendo particular relevância económica a espécie Synchytrium endobioticum (Schilb.) Percival (1909) (Chytridiales) é responsável pela doença conhecida como verruga-preta-da-batata.[28]
Também em relação aos animais, algumas espécies são parasitas agressivos, conhecendo-se uma capaz de provocar a morte de anfíbios em larga escala. O processo, no entanto, é desconhecido. A infeção é denominada quitridiomicose. O declínio das populações de anfíbios levou à descoberta desta infeção em 1998, na Austrália e no Panamá. Espécies no género Batrachochytrium são responsáveis por declínios severos nas populações de anfíbios globalmente.[29]
Batrachochytrium dendrobatidis[editar | editar código-fonte]
.jpg)
A espécie Batrachochytrium dendrobatidis tem merecido especial atenção por ser considerada responsável pelo declínio da população de anfíbios nativos em várias partes do mundo, inclusive no Brasil.[30] A espécie é responsável pela quitridiomicose, uma grave doença em diversos grupos de animais, mas particularmente mortífera quando ataca anfíbios. Descoberto em 1998 na Austrália e Panamá, esta doença é conhecida por matar anfíbios em grande número, e tem sido apontada como a principal causa para o declínio das populações de anfíbios à escala global.
Surtos deste fungo foram considerados responsáveis por matar grande parte da população de Nectophrynoides asperginis (o sapo-de-kihansi) no seu habitat nativo da Tanzânia,[31] bem como pela extinção da espécie Incilius periglenes (o sapo-dourado) em 1989. A quitridiomicose também foi implicada na suposta extinção de Rheobatrachus silus (a rã-gástrica-do-sul),[32] vista pela última vez na natureza em 1981, e da Rheobatrachus eungellensis (a rã-gástrica-do-norte), registada pela última vez na natureza em março de 1985.[33]
Acredita-se que o processo que leva à mortalidade dos anfíbios seja a perda de iões essenciais através dos poros produzidos nas células epidérmicas pelo quitrídio durante a sua replicação.[34] Investigação recente revelou que elevar ligeiramente os níveis de sal nas águas pode curar a quitridiomicose em algumas espécies de sapos australianos,[35] embora seja necessária mais experimentação para confirmar a viabilidade desta potencial abordagem ao controlo da quitridiomicose em anfíbios.
Classificação[editar | editar código-fonte]
Tal como em relação a outros grupos de organismos, as espécies de Chytridiomycota têm sido tradicionalmente delineadas e classificadas com base no desenvolvimento, morfologia, substrato e método de descarga dos zoósporos.[14][9] No entanto, mesmo cultura realizadas a partir de isolados de esporos únicos (ou linhas isogénicas) exibem uma grande quantidade de variação em muitas dessas características, pelo que essas características não podem ser usadas para classificar ou identificar uma espécie de forma confiável.[14][9][36] Atualmente, a taxonomia dos Chytridiomycota é baseada em dados moleculares, ultraestrutura dos zoósporos e alguns aspectos da morfologia e desenvolvimento do talo.[14][36]
Taxonomia[editar | editar código-fonte]
Em um sentido mais antigo e restrito (não usado aqui), o termo quitrídio (ou por vezes quitrídia) refere apenas os fungos da classe Chytridiomycetes. Aqui, o termo "quitrídio" refere-se a todos os membros de Chytridiomycota.[4] Em tempos ainda mais antigos, os quitrídios foram incluídos entre os Protoctista,[14] mas agora são regularmente classificados como fungos.
Em classificações mais antigas, os quitrídios, exceto os recentemente incluídos na ordem Spizellomycetales, foram colocados na classe Phycomycetes sob o subfilo Myxomycophyta do reino Fungos. Anteriormente, estiveram colocados no grupo Mastigomycotina como a classe Chytridiomycetes.[37] As outras classes de Mastigomycotina, os Hyphochytriomycetes e oomycetes, foram retiradas dos fungos para serem classificadas como heterocontes pseudofúngicos.[38]
Na sua presente circunscrição taxonómica, a classe Chytridiomycetes agrupa mais de 750 espécies de quitrídios distribuídas por 13 ordens.[12][39][40][41] Antes da revisão de 2018, o grupo incluía as classes adicionais Monoblepharidomycetes,[42] com duas ordens, e Hyaloraphidiomycetes com uma única ordem.[43] Aquela revisão elevou aquelas classes ao nível taxonómico de divisão (ou filo), como Monoblepharomycota (onde se incluiu a classe Hyaloraphidiomycetes), sendo-lhe adicionadas as divisões Caulochytriomycota e Neocallimastigomycota (sinónimos taxonómicos pro parte da classe original), passando o conjunto a formar o clado Chytridiomyceta, geralmente considerado ao nível taxonómico de sub-reino.[12]
A aplicação das técnicas da filogenia molecular, em combinação com outras técnicas, como a análise ultraestrutural, aumentou muito a compreensão da filogenia dos quitrídios e levou à formação de vários novos filos de fungos zoospóricos:
- Os membros da ordem Blastocladiales, originalmente parte dos Chytridiomycota, são agora classificados como um filo separado, a divisão Blastocladiomycota.[44]
- Os membros da ordem Neocallimastigales, originalmente uma ordem de fungos anaeróbicos da classe Chytridiomycetes, encontrada no trato digestivo de herbívoros, foi elevada a um filo separado, a divisão Neocallimastigomycota.[43]
- Os membros da família Olpidiaceae, incluindo o género tipo Olpidium, anteriormente classificados na ordem Chytridiales, foram elevados a um filo separado, a divisão Olpidiomycota.[45]
Filogenia[editar | editar código-fonte]
As características deste grupo situam-se evolutivamente como um grado intermediário entre os fungos primitivos unicelulares (Aphelidiomycota e Rozellomycota) e os fungos terrestres mais evoluídos (Amastigomycota) cujos esporos não apresentam flagelos.
Devido a ser comum o modo de vida saprófito nos quitrídios, a principal tendência evolutiva pode ter sido o aumento da capacidade de degradar a matéria orgânica indigerível para outros seres vivos, em particular a lignocelulose (biomassa de lignina e celulose ou hemicelulose) e é possível que esta característica tenha sido suportada na radiação evolutiva dos fungos mais avançados.[46]
A natureza do esporo uniflagelado, compartilhada com Aphelidiomycota e Rozellomycota, paralelamente ao que ocorre com os espermatozóides uniflagelados em animais, bem como a análise molecular, cria o traço evolutivo que relaciona estes organismos com os reinos Fungi e Animalia e com o clado eucariótico Opisthokonta, onde ancestralmente os protozoários tipicamente possuíam um único flagelo inserido posteriormente.
Os quitrídios na classificação tradicional eram um táxon parafilético porque Blastocladiomycota (anteriormente Blastocladiales) está mais intimamente relacionado com os fungos terrestres (Amastigomycota) do que com os outros quitrídios. Foi proposta uma nova classificação filogenética, baseada em critérios que permitem criar agrupamentos monofiléticos, que reparte os quitrídios por múltiplas divisões.[47][12] Uma possível árvore filogenética é a seguinte:[48][49][50]
| Fungi |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Registo fóssil[editar | editar código-fonte]
Os fósseis mais antigos de quitrídios que se conhecem são da formação Rhynie chert (Escócia), uma lagerstätte do Devoniano com preservação anatómica de plantas e fungos. Entre os microfósseis estão quitrídios preservados como parasitas em riniófitos (†Rhyniophytes). Esses fósseis muito semelhantes ao género moderno Allomyces.[51]
Restos de quitrídios holocárpicos foram encontrados em cherts de Combres, no centro da França, que datam do final Viseano. Esses restos foram encontrados junto com restos eucárpicos e são de natureza ambígua, embora se pense que sejam de quitrídios.[52] Outros fósseis semelhantes a quitrídios foram encontrados em sílex do Pensilvaniano na Bacia de Saint-Etienne, em França, datando entre 300~350 Ma.[53]
Uso em ficção[editar | editar código-fonte]
A novela Tom Clancy's Splinter Cell: Fallout (2007) apresenta uma espécie de quitrídio que se alimenta de petróleo e derivados de petróleo. Na história, a espécie é modificada usando radiação nuclear, para aumentar a taxa na qual se alimenta de óleo. É então usado por extremistas islâmicos na tentativa de destruir o abastecimento mundial de petróleo, tirando assim a vantagem tecnológica aos Estados Unidos.[54]
O webcomic Black Mudpuppy, de Ethan Kocak, apresenta um arco com um quitrídio antropomórfico maligno desenvolvido como uma experiência nazi para infectar Xolotl, um axolotl negro gigante capaz de falar.[55]
Referências[editar | editar código-fonte]
- ↑ a b Tedersoo, Sánchez-Ramírez, Kõljalg, Bahram, M.Döring, Schigel, T.W.May, M.Ryberg & Abarenkov, Fungal Diversity 90(1): 148 (2018). [Index Fungorum ID: 554033]
- ↑ Hassett, B.T.; Gradinger, R. (2016). «Chytrids dominate arctic marine fungal communities». Environ Microbiol. 18 (6): 2001–2009. PMID 26754171. doi:10.1111/1462-2920.13216
- ↑ Kilias, Estelle S.; Junges, Leandro; Šupraha, Luka; Leonard, Guy; Metfies, Katja; Richards, Thomas A. (2020). «Chytrid fungi distribution and co-occurrence with diatoms correlate with sea ice melt in the Arctic Ocean». Communications Biology. 3 (1): 183. PMC 7174370
 . PMID 32317738. doi:10.1038/s42003-020-0891-7
. PMID 32317738. doi:10.1038/s42003-020-0891-7
- ↑ a b c Alexopoulos, C.J.; Mims, C.W.; Blackwell, M. (1996). Introductory Mycology 4th ed. [S.l.]: John Wiley & Sons
- ↑ a b c d e Kendrick, Bryce (2000). The Fifth Kingdom 3rd ed. Newburyport, MA: Focus Publishing
- ↑ a b Zottarell, Carmen Lidia Amorim Pires. «Contribuição para o conhecimento de Chytridiomycota da "Reserva Biológica de Paranapiacaba", Santo André, SP, Brasil»
- ↑ James, Timothy Y. «A molecular phylogeny of the flagellated fungi (Chytridiomycota) and description of a new phylum (Blastocladiomycota)»
- ↑ Tedersoo, L., Sánchez-Ramírez, S., Kõljalg, U., Bahram, M., Döring, M., Schigel, D.S., May, T.W., Ryberg, M. & Abarenkov, K. 2018. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Diversity 90(1): 135–159. DOI: 10.1007/s13225-018-0401-0.
- ↑ a b c d e f g h i j k l m n o Sparrow, F.K. (1960). Aquatic Phycomyete 2nd ed. Ann Arbor, MI: The University of Michigan Press
- ↑ Blackwell, M. (2011). «The Fungi: 1,2,3 … million species?». American Journal of Botany. 98 (3): 426–438. PMID 21613136. doi:10.3732/ajb.1000298
- ↑ Longcore, J.E.; Pessier, A.P.; Nichols, D.K. (1999). «Batrachochytirum dendrobatidis gen. et sp. nov., a chytrid pathogenic to amphibians». Mycologia. 91 (2): 219–227. doi:10.1080/00275514.1999.12061011
- ↑ a b c d
Tedersoo, Leho; Sanchez-Ramırez, Santiago; Koljalg, Urmas; Bahram, Mohammad; Doring, Markus; Schigel, Dmitry; et al. (22 fevereiro 2018). «High-level classification of the Fungi and a tool for evolutionary ecological analyses». Fungal Diversity. 90 (1): 135–159. doi:10.1007/s13225-018-0401-0
- ↑ a b Alexopoulos, C.J., Mims, C.W. & Blackwell, M.B., Introductory Mycology. 4th ed., John Wiley & Sons, New York, 1996.
- ↑ a b c d e f g h i j Barr, D.J.S. (1990). Margulis; Corliss; Melkonian; Chapman, eds. Handbook of Protoctista. Boston, MA: Jones & Barlett. pp. 454–466
- ↑ Hawksworth, D.L.. Kirk, P.M., Sutton, B.C. & Pegler, D.M. 1995. Ainsworth & Bisby´s Dictionary of the Fungi. 8th ed. International Mycological Institute, Egham.
- ↑ Raven, Peter H., Biologia Vegetal / Peter . Raven, Ray F. Evert, Susan E. Eichhorn; Rio de Janeiro : Guanabara Koogan, 2010.
- ↑ Hibbett; et al. (2007). «A higher-level phylogenetic classification of the Fungi». Mycologia. 111 (5): 509–547. PMID 17572334. doi:10.1016/j.mycres.2007.03.004
- ↑ Powell MJ, Chambers JG, Holznagel WE. 2004b. Phylogenetic relationships among Rhizophydium isolates from North America and Australia. Mycologia 96: 1339–1351.
- ↑ Barr, D. J. S. (2001). Chytridiomycota. In D. J. McLaughlin, E. G. McLaughlin, & P. A. Lemke (Eds.), The Mycota (Vol. 7A, pp. 93–112). New York: Springer.
- ↑ «The Chytridiomycota». Mycology. nbm-mnb.ca. Natural History of Fungi. Consultado em 28 de outubro de 2013
- ↑ Carlile, M.J. (1986). Ayres, Peter G.; Boddy, Lynne, eds. Water, Fungi, and Plants. 11. [S.l.]: Cambridge University Press
- ↑ Letcher, P.M.; et al. (2008). «Rhizophlyctidales – a new order in Chytridiomycota». Mycological Research. 112 (9): 1031–1048. PMID 18701267. doi:10.1016/j.mycres.2008.03.007
- ↑ Simmons, D.R. (2011). «Phylogeny of Powellomycetacea fam. nov. and description of Geranomyces variabilis gen. et comb. nov». Mycologia. 103 (6): 1411–1420. PMID 21558503. doi:10.3852/11-039
- ↑ Lefèvre, E.; Letcher, P.M.; Powell, M.J. (2012). «Temporal variation of the small eukaryotic community in two freshwater lakes: Emphasis on zoosporic fungi». Aquatic Microbial Ecology. 67 (2): 91–105. doi:10.3354/ame01592
- ↑ Freeman, K.R. (2009). «Evidence that chytrids dominate fungal communities in high-elevation soils». Proceedings of the National Academy of Sciences of the USA. 106 (43): 18315–18320. Bibcode:2009PNAS..10618315F. PMC 2775327. PMID 19826082. doi:10.1073/pnas.0907303106
- ↑ Ibelings, B.W.; de Bruin, A.; Kagami, M.; Rijkeboer, M.; van Donk, E. (2004). «Host parasite interactions between freshwater phytolankton and chytrid fungi (chytridiomycota)». J. Phycol. 40 (3): 457–455. doi:10.1111/j.1529-8817.2004.03117.x
- ↑ Gleason, Frank H.; et al. (2008). «The ecology of chytrids in aquatic ecosystems: roles in food web dynamics». Fungal Biology Reviews. 22 (1): 17–25. doi:10.1016/j.fbr.2008.02.001
- ↑ Hooker, W.J. (1981). Compendium of Potato Diseases. [S.l.]: International Potato Center. pp. 36–7. ISBN 978-0-89054-027-5
- ↑ Recent Asian origin of chytrid fungi causing global amphibian declines por Simon J. O’Hanlon et al Vol. 360, edição 6389, pp. 621-627 DOI: 10.1126 / science.aar1965 (2018)
- ↑ Carnaval, A.C.O.Q., Toledo, L.F., Haddad, C.F.B. & Britto, F.B. 2005. Chytrid fungus infects high-altitude stream-dwelling Hylodes magalhaesi (Leptodactylidae) in the Brazilian Atlantic rainforest. Froglog 70:3.
- ↑ Dean, Cornelia (1 fevereiro 2010). «Saving tiny toads without a home». The New York Times
- ↑ Rheobatrachus silus species. Environment (Relatório). Profile and threats database. Government of Australia
- ↑ Rheobatrachus eungellensis species. Environment (Relatório). Profile and threats database. Government of Australia
- ↑ Voyles, J.; L. Berger; S. Young; et al. (2007). «Electrolyte depletion and osmotic imbalance in amphibians with chytridiomycosis». Diseases of Aquatic Organisms. 77 (2): 113–118. PMID 17972752. doi:10.3354/dao01838
- ↑ «A salty cure for a deadly frog disease». Australian Broadcasting Corporation. 5 de fevereiro de 2018
- ↑ a b Blackwell, W.H.; Letcher, P.M.; Powell, M.J. (2006). «Thallus development and the systematics of Chytridiomycota: an additional developmental pattern represented by Podochytrium». Mycotaxon. 97: 91–109
- ↑ Ainsworth; Sparrow; Sussman, eds. (1973). The Fungi: An advanced treatise. IVB A – Taxonomic review with keys: Basidiomycetes and lower fungi. New York, NY: Academic Press
- ↑ van der Auwera, G.; de Baere, R.; van de Peer, Y.; de Rijk, P.; van den Broeck, I.; de Wachter, R. (Julho 1995). «The phylogeny of the Hyphochytriomycota as deduced from ribosomal RNA sequences of Hyphochytrium catenoides». Mol. Biol. Evol. 12 (4): 671–678. PMID 7659021. doi:10.1093/oxfordjournals.molbev.a040245
- ↑ «Chytrid fungi online – an NSF / PEET project website». Tuscaloosa, AL: University of Alabama. Consultado em 16 de abril de 2022
- ↑ Esser, K. (2014). The Mycota. VII A: Systematics and evolution 2nd ed. [S.l.]: Springer. p. 461. ISBN 978-3-642-55317-2
- ↑ Powell; Letcher (20 janeiro 2017) [2015]. «A new genus and family for the misclassified chytrid, Rhizophlyctis harderi». Mycologia. 107 (2): 419–431. PMID 25572098. doi:10.3852/14-223. Consultado em 23 de agosto de 2016
- ↑ Dee, J.M.; Mollicone, M.; Longcore, J.E.; Roberson, R.W.; Berbee, M.L. (2015). «Cytology and molecular phylogenetics of Monoblepharidomycetes provide evidence for multiple independent origins of the hyphal habit in the Fungi». Mycologia. 107 (4): 710–728. ISSN 0027-5514. PMID 25911696. doi:10.3852/14-275
- ↑ a b Misra, J.K.; Tewari, J.P.; Deshmukh, S.K. (10 janeiro 2012). Systematics and Evolution of Fungi. [S.l.]: CRC Press. p. 47. ISBN 978-1-57808-723-5
- ↑ James, T.Y.; et al. (2006). «A molecular phylogeny of the flagellated fungi (Chytridiomycota) and description of a new phylum (Blastocladiomycota)». Mycologia. 98 (6): 860–871. PMID 17486963. doi:10.3852/mycologia.98.6.860
- ↑ «Item no. 42» (PDF). Index Fungorum
- ↑ Philippe Silar 2016, Protistes Eucaryotes.: Origine, Evolution et Biologie des Microbes Eucaryotes, 2016, 978-2-9555841-0-1. <hal-01263138>
- ↑ Miguel A. Naranjo‐Ortiz et Toni Gabaldón. Fungal evolution: diversity, taxonomy and phylogeny of the Fungi Wiley Online Library.
- ↑ «A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data». ISSN 0027-5514. PMC 6078412. PMID 27738200. doi:10.3852/16-042
- ↑ Yuanning Li, Jacob L Steenwyk, Ying Chang, Yan Wang, Timothy Y James, Jason E Stajich, Joseph W Spatafora, Marizeth Groenewald, Casey W Dunn, Chris Todd Hittinger, Xing-Xing Shen, Antonis Rokas (2021). A genome-scale phylogeny of the kingdom Fungi. Cell.com.
- ↑ Tedersoo, L., Sánchez-Ramírez, S., Kõljalg, U. et al. 2018, High-level classification of the Fungi and a tool for evolutionary ecological analyses Fungal Diversity 90: 135. https://doi.org/10.1007/s13225-018-0401-0
- ↑ Taylor, T.N.; Remy, W.; Hass, H. (1994). «Allomyces in the Devonian». Nature. 367 (6464): 601. Bibcode:1994Natur.367..601T. doi:10.1038/367601a0
- ↑ Krings, Michael; Dotzler, Nora; Taylor, Thomas; Galtier, Jean (2009). «Microfungi from the upper Visean (Mississippian) of central France: Chytridiomycota and chytrid-like remains of uncertain affinity». Review of Palaeobotany and Palynology. 156 (3–4): 319–328. doi:10.1016/j.revpalbo.2009.03.011
- ↑ Krings, Michael; Jean Galtier; Thomas N. Taylor; Nora Dotzler (2009). «Chytrid-like microfungi in Biscalitheca cf. musata (Zygopteridales) from the Upper Pennsylvanian Grand-Croix cherts (Saint-Etienne Basin, France)». Review of Palaeobotany and Palynology. 157 (3–4): 309–316. doi:10.1016/j.revpalbo.2009.06.001
- ↑ Michaels, David (2007). Tom Clancy's Splinter Cell: Fallout. [S.l.]: Penguin Group. ISBN 978-0-425-21824-2
- ↑ «Black Mudpuppy». Chytrid crisis: Part 1. 17 de outubro de 2016. Consultado em 29 de agosto de 2017
Ligações externas[editar | editar código-fonte]
- «Chytrid fungi online – an NSF / PEET project website». Tuscaloosa, AL: University of Alabama. Consultado em 16 de abril de 2022
- «Longcore Lab». Orono, ME: University of Maine. Cópia arquivada em 13 de dezembro de 2013 — Includes links on how to isolate and culture chytrids.
- «Introduction to the Chytridiomycota». U.C. Museum of Paleontology. Berkeley, CA: University of California, Berkeley
- «Impact of chytrid fungus on frogs». Australia: Foundation for National Parks & Wildlife. Cópia arquivada em 10 de janeiro de 2007
- «'Amphibian Ark' aims to save frogs from fungus». NBC News
- Chytrid fungus and how it impacts amphibians kept as domestic pets in the Aquarium Wiki Encyclopaedia.
- MeSH Chytridiomycota