A história genética da Europa é a área da arqueogenética que estuda a formação, etnogênese e outras informações genéticas específicas sobre as populações nativas da Europa.
A dispersão principal de humanos para fora da África, ocorrida há cerca de 70 mil anos, deu origem a uma linhagem distinta da dos africanos. Há cerca de 50 000 anos esta linhagem primitiva ter-se-á dividido nas linhagens da Eurásia Oriental e Ocidental.[1][2][3][4] Todos os humanos que habitam fora da África possuem um pouco de genética dos neandertais.[5]
Como resultado das migrações ocorridas entre o Mesolítico e a Idade do Bronze, as populações nativas da Europa modernas se distinguem entre si por possuírem diferentes porcentagens de ancestralidade WHG, EEF e ANE.[11][12][13] As taxas de miscigenação variaram de acordo com o local: no final do Neolítico, a ancestralidade WHG em agricultores na Hungria era de aproximadamente 10%, na Alemanha era de cerca de 25% e na Península Ibérica, tão alta quanto 50%.[11] A contribuição genética dos EEF é mais significativa na Europa Mediterrânea e diminui em direção ao norte e nordeste da Europa, onde a ascendência WHG é mais forte; os sardos são a etnia europeia mais próxima dos primeiros agricultores europeus. A ascendência ANE é encontrada em toda a Europa em porcentagens reduzidas, com máximos de cerca de 20% encontrados em povos bálticos e finlandeses.
A pesquisa sobre a história genética da Europa tornou-se possível na segunda metade do século XX, mas só produziu resultados contundentes preliminares na década de 1990, ainda restritos a estudos dos haplogrupos do DNA mitocondrial e cromossomo Y. O estudo do DNA autossômico tornou-se mais acessível na década de 2000 e, desde meados da década de 2010, resultados de resolução anteriormente inatingível, muitos deles baseados em análises dos genomas antigos completos, foram publicados em ritmo acelerado.[14][15]
Devido à seleção natural, a porcentagem de DNA neandertal nos europeus arcaicos foi diminuindo gradualmente com o passar dos milênios. Há entre 45 mil anos e 7 mil anos, a porcentagem caiu de cerca de 3% a 6% para 2%.[15] A remoção de alelos neandertais ocorreu com mais frequência em torno dos genes do que em outras partes do genoma.[15]
Distribuição do Homem de Neandertal, com os principais sítios arqueológicos em que foram encontrados fósseis dessa espécie.
Os neandertais habitavam grande parte da Europa, Oriente Médio e Ásia Central há centenas de milhares de anos, existindo no continente europeu até aproximadamente há 30 mil anos. Eventualmente, foram substituídos pelos humanos anatomicamente modernos, que chegaram pela primeira vez à Europa há cerca de 40 mil anos. Haja vista que as duas espécies de hominídeos provavelmente coexistiram na Europa, antropólogos e arqueólogos há muito perguntam se ambas as espécies interagiram.[16][17] Tal questão foi solucionada apenas em 2010, quando foi descoberto que as populações da Eurásia apresentam, em média, uma porcentagem de 1,5-2,1% de genes neandertais.[18] A partir de então, a dúvida passaria a ser se essa miscigenação ocorreu na Europa ou no Levante.
Também houve especulações sobre genes específicos que teriam sido herdados dos neandertais. Por exemplo, um locus MAPT17q 21.3, dividido em linhagens genéticas profundas H1 e H2. Como a linhagem H2 aparenta ser restrita às populações europeias, vários pesquisadores argumentaram que esta é uma herança neandertal, o que começou em 2005.[19][20][21][22] Entretanto, os resultados preliminares do sequenciamento completo do material genético dos neandertais nos anos 2000 não conseguiram detectar evidências de cruzamentos com humanos modernos.[23][24] Em 2010, as descobertas de Svante Pääbo (Instituto Max Planck de Antropologia Evolutiva), Richard E. Green (Universidade da Califórnia em Santa Cruz) e David Reich (Escola de Medicina Harvard), feitas por meio da comparação do material genético dos ossos de três neandertais com o de cinco humanos modernos, mostraram uma relação entre neandertais e humanos modernos fora da África.
Expansão dos humanos modernos para fora da África e substituição dos neandertais
Acredita-se que os humanos modernos começaram a habitar a Europa a partir de há 40 mil anos, no Paleolítico Superior. Algumas evidências mostram a disseminação da cultura Aurignaciana.[25]
De uma perspectiva dos haplogrupos do cromossomo Y, possivelmente os antigos haplogrupos C1a2, F e/ou E são aqueles cuja presença na Europa é mais antiga, visto que foram encontrados em alguns restos humanos bastante antigos. Entretanto, outros haplogrupos são muito mais comuns entre os homens europeus modernos devido às mudanças demográficas posteriores.
O haplogrupo I (M170), atualmente relativamente comum e difundido na Europa, pode representar um marcador do Paleolítico, pois sua idade foi estimada em 22 mil anos. Embora hoje esteja concentrado na Europa, provavelmente surgiu na região do Oriente Médio ou do Cáucaso, quando divergiu de seu ancestral imediato, o haplogrupo IJ. Nessa mesma época, também surgiu a Cultura Gravetiana.[26]
Pesquisas mais antigas sobre o DNA do cromossomo Y se concentram no haplogrupo R1 (M173), o mais numeroso entre os homens europeus atuais. Acredita-se que tenha surgido há aproximadamente 40 mil anos na Ásia Central.[26][27] Apesar disso, atualmente estima-se que R1 surgiu mais recentemente: um estudo de 2008 datou ancestral comum mais recente do haplogrupo IJ há 38500 anos e do haplogrupo R1 há 18 mil anos. Isso sugere que os colonos do haplogrupo IJ formaram a primeira onda e o haplogrupo R1 chegou muito mais tarde.[28]
Dessa forma, os dados genéticos sugerem que, do ponto de vista da ascendência patrilinear, grupos de humanos modernos distintos tomaram duas rotas distintas para a Europa: uma, da região do Oriente Próximo para os Balcãs e outra, da Ásia Central pela estepe eurasiática.
Foi descoberto por Martin Richards et al. que entre 15 e 40% das linhagens de DNA mitocondrial existentes remontam às migrações paleolíticas, o que depende da existência de vários eventos fundadores.[29] O haplogrupo mitocondrial U5, datado de há 40 a 50 mil anos, chegou ao continente durante a primeira onda colonização. ainda no Paleolítico Superior, sendo responsável por 5 a 15% do total de linhagens do DNA mitocondrial. Os movimentos do UP médio são marcados pelos haplogrupos HV, I e U4. HV dividiu-se em Pré-V (cerca de 26 mil anos) e o ramo maior H, ambos dispersos pela Europa, possivelmente através de contatos durante o Gravetiano.[26][30]
O Haplogrupo H do DNA mitocondrial é responsável por aproximadamente metade das linhas de genes na Europa, com muitos subgrupos. As linhagens mitocondriais acima ou seus percussores chegaram teriam chegado à Europa através do Oriente Médio. Isso contrasta com a evidência do cromossomo Y, visto que em torno de 50% das linhagens masculinas são caracterizadas pela superfamília R1, oriundas provavelmente da Ásia Central. Ornella Semino postula que essas diferenças "podem ser parcialmente devido à idade molecular aparente mais recente dos cromossomos Y em relação a outros loci, o que sugere uma substituição mais rápidas dos haplogrupos anteriores de cromossomo Y. Migrações diferenciadas baseadas em gênero também influenciaram os padrões observados em variação dos haplogrupos do DNA mitocondrial e cromossomo Y.
Este fato diminuiu a diversidade genética na Europa, sendo um "resultado da deriva, consistente com um gargalo populacional inferido durante o Último Máximo Glacial".[27] Ao passo que as geleiras derretiam há entre 16 e 13 mil anos, as regiões mais ao norte da Europa foram sendo gradualmente repovoadas por pessoas vindas das regiões de refúgio mais ao sul.[26]
Alguns clados do haplogrupo I do cromossomo Y aparentam ter se divergido se seus haplogrupos parentais em algum momento durante ou logo após o Último Máximo Glacial.[32] O haplogrupo I2 é predominante nos Balcãs ocidentais, bem como no restante do sudeste e centro-leste da Europa em frequências mais moderadas. Sua presença cai rapidamente na Europa Central, o que sugere que os sobreviventes com as linhagens I2 se expandiram predominantemente pelo sudeste e centro-leste da Europa.[33]
De uma perspectiva do DNA mitocondrial, Richards et al. descobriram que a maior parte da diversidade dos haplogrupos do DNA mitocondrial na Europa é explicada pelas reexpansões pós-glaciais ocorridas durante o final do Paleolítico Superior e o Mesolítico. "As análises regionais evidenciam a hipótese de que grande parte da Europa ocidental e central foi repovoada a partir principalmente do sudoeste quando o clima melhorou. As linhagens envolvidas incluem muito do haplogrupo mais comum, H, bem como muito dos haplogrupos K, T, W e X." O estudo não pôde determinar se ocorreram novas migrações de linhagens do DNA mitocondrial do Oriente Próximo durante este período; um entrada significativa foi considerada improvável.[29]
O modelo alternativo de um número maior de refugiados foi discutido por Bilton et al.[34]
A partir da análise de 51 indivíduos, os pesquisadores puderam identificar cinco agrupamentos genéticos separados de europeus arcaicos durante o Último Máximo Glacial: o de Dolní Věstonice (na atual Chéquia, há entre 34 a 25 mil anos), associado à cultura gravetiana; o de Mal'ta dos Antigos eurasiáticos do Norte (Sibéria, 24 a 17 mil anos antes do presente), associado à Cultura Mal'ta-Buret; a Caverna El Mirón (Espanha, 19 a 14 mil anos antes do presente), associada à Cultura Magdaleniana; Ripari Villabruna (Itália, 14 a 7 mil anos antes do presente).[15] o Aglomerado Mal'ta (24.000–17.000), associado à cultura Mal'ta-Buret', o Agrupamento El Mirón (19.000–14.000 anos antes do presente), associado à cultura Magdaleniana ; o Cluster Villabruna (14.000-7.000 anos antes do presente) e o Cluster Satsurblia (13.000 a 10.000 anos antes do presente).[15]
A partir de há cerca de 37 mil anos, todos os europeus arcaicos passaram a compartilhar alguma ancestralidade com os europeus modernos.[15] Esta população fundadora é representada pelo espécime GoyetQ116-1, de 35 mil anos da Bélgica.[15] Esta linhagem desaparece do registro fóssil e não é encontrada novamente até há 19 mil anos na Caverna de El Mirón, cujos indivíduos mostram fortes afinidades com GoyetQ116-1.[15] Durante este intervalo, o distinto agrupamento de Dolni-Věstonice é predominante na Europa, mesmo em Goyet.[15] A reexpansão do Aglomerado da Caverna El Mirón coincidiu com o aumento das temperaturas após o recuo das geleiras durante o Último Máximo Glacial.[15] Há entre 37 a 14 mil anos, a população da Europa consistia em um grupo isolado descendente de uma população fundadora que não cruzou significativamente com outras populações.[35]
As populações do Mesolítico divergiram significativamente, por causa do seu relativo isolamento ao longo de muitos milênios, às duras pressões seletivas durante o Último Máximo Glacial e aos efeitos fundadores causados pela rápida expansão no Mesolítico a partir dos refúgios glaciais.[6] No final do Último Máximo Glacial, há entre aproximadamente 19 a 11 mil anos, surgiram as variedades familiares de fenótipos eurasianos. Os Caçadores-coletores ocidentais (WHG em inglês), que habitavam a Europa Ocidental e Meridional e a maioria da Europa Central, apresentavam pele escura, herdada dos primeiros europeus antes do Último Máximo Glacial, e olhos claros[36] e sua contribuição genética não é majoritária em nenhuma população europeia moderna.[37] As variações HERC2 e OCA2 para olhos azuis são derivadas da linhagem WHG e também foram encontradas nos proto-indo-europeus.[37]
Há aproximadamente 14 mil anos, o aglomerado de Ripari Villabruna afastou-se da afinidade com o GoyetQ116-1 e começou a mostrar mais afinidade com o Oriente Próximo, uma mudança que coincide com o aumento das temperaturas do interestágio Bølling-Allerød.[15] Essa mudança genética mostra que as populações do Oriente Próximo teriam começaram a se mudar para a Europa durante o final do Paleolítico Superior, aproximadamente 6 mil anos antes do que se pensava anteriormente, antes da introdução da agricultura.[35] Alguns espécimes do Aglomerado de Ripari Villabruna também mostram afinidades genéticas com os asiáticos orientais que são derivadas do fluxo gênico.[15][35] A variação HERC2 para olhos azuis apareceu pela primeira vez há aproximadamente 14 mil anos na Itália e no Cáucaso.[15]
Com o fim do Último máximo glacial, há cerca de 14 mil anos, grupos de Caçadores-Coletores Ocidentais deixaram os refúgios glaciais e migraram para o restante da Europa. No Leste Europeu e Escandinávia, eles se encontram e misturaram com os Antigos Eurasiáticos do Norte (ANE em inglês)[36], de pele clara e majoritariamente loiros[9], oriundos da Sibéria e da Ásia Central. Dessa mistura, surgiram os Caçadores-Coletores Orientais (EHG em inglês) e os Caçadores-Coletores Escandinavos (SHG em inglês), ambos de pele branca e com cores de olhos e cabelos variáveis.[36] Os EHG possuíam em torno de três quartos de sua ancestralidade oriunda dos ANE, enquanto os SHG possuíam menos dessa linhagem - entre 19 e 37%.[10][38][39]
Os ANE são associados à Cultura Mal'ta-Buret da Sibéria[40] e seus últimos indivíduos viveram na Ásia Central e Xinjiang há cerca de três mil anos.[9][41] De acordo com Lazaridis, "a ancestralidade dos Antigos Eurasiáticos do Norte é proporcionalmente o menor componente em toda a Europa, nunca maior que 20%, mas encontrado em quase todos os grupos europeus estudados".[42] Esse componente genético não vem diretamente dos indivíduos de Mal'ta em si, mas de uma linhagem relacionada que se separou da linhagem Mal'ta.[15]
O alelo derivado do gene KITLG (SNP rs12821256), que está associado e provavelmente é causal para os cabelos loiros em europeus, foi encontrado nos caçadores-coletores da Europa Oriental e da Escandinávia, mas não nos da Europa Ocidental, o que sugere que os cabelos loiros foram introduzidos na Europa pelos Antigos Eurasiáticos do Norte e se espalharam pelo continente graças às migrações indo-europeias, de indivíduos vindos da Estepe Pôntica, os quais parcialmente descendiam dos ANE. Consistente com isso, o mais antigo indivíduo conhecido com o alelo derivado para cabelos loiros era dos Antigos Eurasiáticos do Norte e foi encontrado no complexo arqueológico de Afontova Gora, na Sibéria central, tendo vivido há 18 mil anos.[43]
Uma grande variação genética na Europa, reconhecida há muito tempo, parece mostrar importantes dispersões vindas do Oriente Próximo. Este fato tem sido frequentemente associado à disseminação da agricultura durante o Neolítico, período que tem sido considerado um dos mais significativos na determinação da diversidade genética da Europa de hoje.
Durante muito tempo, os estudiosos pensavam que a agricultura chegou à Europa por intermédio de mercadores, não por migrantes.[44] No entanto, com os estudos genéticos da população europeia, foi descoberto que a agricultura foi introduzida no continente por meio das migrações de agricultores da Anatólia, os primeiros agricultores europeus (EEF em inglês), os quais tinham a pele clara e majoritariamente cabelos e olhos escuros.[36]
Os primeiros agricultores europeus não eram descendentes dos que descobriram a agricultura no Crescente Fértil há 12 mil anos, mas sim de caçadores-coletores da própria Anatólia que adotaram o estilo de vida de seus vizinhos.[45]
Mapa da dispersão da agricultura pela Europa a partir do Oriente Próximo. Os números indicam anos antes de Cristo.
A introdução da agricultura na Europa levou a gigantescas mudanças econômicas e socioculturais no continente, o que também gerou um grande efeito na sua diversidade genética, especialmente no tocante às linhagens genéticas que entram na Europa do Oriente Médio para os Balcãs. Houve várias fases deste período:
Em um prelúdio do Mesolítico tardio para o neolítico, aparentemente os povos do Oriente Próximo de áreas que já tinham agricultura e que também possuíam tecnologia marítima tiveram uma presença transitória na Grécia (por exemplo, na caverna de Franchthi).[46][47]
Há um consenso de que a agricultura e sua tecnologia e as principais espécies de animais e plantas cultivadas na Europa entraram no continente vindos de alguma parte do Crescente Fértil.[25]:1143, 1150.[48] Menos certamente, algumas vezes argumenta-se que essa revolução agrícola foi parcialmente desencadeada por movimentos de pessoas e tecnologia vindos da África através do Sinai.
Esquema simplificado da história demográfica da Europa durante o Neolítico, com a introdução da agricultura vinda do Oriente Próximo.[49]Em uma fase posterior do Neolítico, o chamado "Neolítico Cerâmico", observou-se a introdução de cerâmica no Levante, Balcãs e sul da Itália (estava presente na região do atual Sudão por algum tempo antes de ser encontrada no Mediterrâneo Oriental, mas acredita-se que tenha se desenvolvido de forma independente), e isso também pode ter sido um período de transferência cultural do Levante para os Balcãs.
Martin Richards estimou que apenas 11% do DNA mitocondrial dos europeus se deve à migração neste período, o que sugere que a agricultura se espalhou principalmente devido à adoção por populações autóctones europeias mesolíticas, e não devido à imigração do Oriente Próximo. O fluxo gênico do Sudoeste para o noroeste da Europa parece ter continuado durante o Neolítico, com a porcentagem declinando significativamente em direção às Ilhas Britânicas. A genética clássica também sugeriu que a maior mistura ao estoque genético europeu do Paleolítico/Mesolítico foi devido à Revolução Neolítica, entre o sétimo e quinto milênio antes de Cristo.[50] Três principais grupos de genes de DNA mitocondrial foram identificados como contribuintes neolíticos para a Europa: em ordem de importância, J, T1 e U3. Com outros, eles representam cerca de 20% do pool genético.[29]
Em 2000, o estudo de Semino sobre o DNA do cromossomo Y revelou a presença de haplótipos pertencentes ao grande clado E1b1b1 (E-M35), encontrados predominantemente no sul dos Balcãs, sul da Itália e partes da Península Ibérica. Semino conectou esse padrão, juntamente com os subclados do haplogrupo J, para ser o componente dos haplogrupos do cromossomo Y da difusão dêmica neolítica de Cavalli-Sforza de agricultores do Oriente Próximo.[26] Rosser et al. observaram um 'componente norte-africano' direto na genealogia europeia, embora não propusessem um momento e um mecanismo para explicá-lo.[51][52] Também descreveu E1b1b como representante de uma migração do Pleistoceno tardio da África para a Europa sobre a Península do Sinai, evidências para as quais não aparecem no DNA mitocondrial.[53]
Entretanto, em relação ao tempo de distribuição e a diversidade do haplogrupo V13, Battaglia (2009)[54] propôs um movimento anterior no qual a linhagem E-M78*, que deu origem a todos os homens com o haplogrupo E-V13, tem origem no sul do Egito e chegou à Europa apenas com as tecnologias mesolíticas. Então, sugerem que o subclado E-V13 de E-M78 expandiu-se apenas posteriormente quando os agricultores-pastores dos Balcãs adotaram tecnologias neolíticas vindas do Oriente Próximo. Eles propõem que a primeira grande dispersão do haplogrupo E-V13 nos Balcãs pode ter sido na direção do Mar Adriático, com a cultura da cerâmica cardial,[33] e propõem que a rota principal da propagação de E-V13 foi ao longo do sistema que abrange as fozes dos rios Vardar, Morava e Danúbio.
Em contraste, Cruciani[55] sugeriu um ponto diferente em que ocorreu a mutação V13 e um tempo de dispersão mais tardio. Os autores propuseram que a mutação V13 apareceu pela primeira vez no Oriente Médio, onde é encontrada em frequências baixas, mas significativas, e entrou nos Balcãs após 11 mil anos antes do Presente. Mais tarde, experimentou uma rápida dispersão que datada em aproximadamente 5300 anos na Europa, coincidindo com a Idade do Bronze dos Balcãs. Assim como Peričic et al., eles consideram que "a dispersão dos haplogrupos E-V13 e J-M12 parece ter seguido principalmente as vias fluviais que ligam o sul dos Balcãs ao centro-norte da Europa".
Mais recentemente, Lacan (2011)[56] anunciou que um esqueleto humano de sete mil anos de idade encontrado em uma caverna da Espanha era um homem com o haplogrupo de cromossomo Y E-V13, enquanto os outros indivíduos testados no mesmo local estavam no haplogrupo G2a, encontrado em restos humanos neolíticos em todo o continente europeu. Usando 7 marcadores STR, este espécime foi identificado como semelhante a indivíduos modernos testados na Albânia, Bósnia e Herzegovina, Grécia, Córsega e Provença. Os cientistas, portanto, propuseram que, se a distribuição moderna de tal haplogrupo do cromossomo Y é resultado de eventos mais recentes, então ele já estava no continente europeu no Neolítico, tendo sido transportado pelos primeiros agricultores europeus vindos do Oriente Próximo. Isso corrobora as propostas de Battaglia et al. em vez de Cruciani et al., pelo menos no que se refere às primeiras dispersões europeias, mas E-V13 pode ter se dispersado mais de uma vez. Ainda mais recente do que a Idade do Bronze, também foi proposto que a distribuição moderna do haplogrupo E-V13 na Europa é pelo menos parcialmente causada por movimentos de pessoas na Roma Antiga.[57]
A migração dos agricultores neolíticos para a Europa trouxe grandes mudanças no continente e várias novas adaptações.[37] No ponto de vista demográfico e natural, a alimentação com produtos vindos da agropecuária aumentou significativamente as taxas de fecundidade, aumentando as taxas de natalidade em relação à Europa pré-agricultura, o que proporcionou um crescimento populacional maior[58]; ao mesmo tempo, zoonoses vieram dos animais domesticados, aumentando as taxas de mortalidade[58], e a necessidade de terras para o plantio e criação de animais levou ao desmatamento, uma prática bastante presente na Europa, que não existia com os caçadores-coletores.[59] Do ponto de vista genético, a variação para a cor da pele clara foi introduzida no continente europeu pelos agricultores vindos do Oriente Próximo,[37] após cuja chegada foi selecionada a mutação SLC22A4, a qual provavelmente surgiu para lidar com a deficiência do aminoácido ergotioneína, mas aumenta o risco de doenças intestinais como a colite ulcerativa, doença celíaca e síndrome do intestino irritável.
A Idade do Bronze presenciou o desenvolvimento de rotas comerciais de longa distância, particularmente ao longo da costa atlântica e no vale do Danúbio. Houve migração da Noruega para os arquipélagos escoceses de Orkney e Shetland neste período e, em menor escala, para a Escócia continental e a Irlanda . Houve também uma migração da Alemanha para o leste da Inglaterra. Martin Richards estima que havia cerca de 4% de imigração de DNA mitocondrial para a Europa na Idade do Bronze.
Sobre a origem e genética dos proto-indo-europeus, esse povo da Estepe Pôntica possuía metade do seu pool genético oriundo dos caçadores-coletores orientais, enquanto a outra metade é oriunda dos caçadores-coletores do Cáucaso (CHG em inglês).[36][40][62]
Os haplogrupos do cromossomo Y R1a e R1b, bastante presentes na Europa, foram dispersos pelo continente pelos proto-indo-europeus, embora esses como um todo possam ser muito mais antigos que a família da linguagem em si.[60][63][64]
No extremo norte, o haplogrupo N do cromossomo Y chegou à Europa vindo da Sibéria com os antigos eurasiáticos do Norte, embora o momento específico de sua chegada seja incerto. É estimado que o subclado N1c1 do norte da Europa mais comum tenha surgido há cerca de 8 mil anos. Há evidências de assentamentos humanos na Finlândia que remontam a 8.500 a.C., ligados à cultura Kunda e seu suposto ancestral, a cultura Swideriana, mas acredita-se que esta última seja de origem europeia. A disseminação do haplogrupo de cromossomo Y N na Europa está fortemente associada à Cultura da Cerâmica de Pente, cujo surgimento é normalmente datado de aproximadamente 4.200 a.C., e com a distribuição das línguas urálicas. Por meio de estudos de DNA mitocondrial dos lapões, o haplogrupo U5 é consistente com múltiplas migrações para a Escandinávia vindas da região entre o Rio Volga e os Montes Urais, iniciadas entre seis e sete mil anos antes do presente.[65]
As relações entre os papéis dos ocupantes vindos da Europa e da Ásia na Finlândia pré-histórica ainda é motivo de discussões e alguns estudiosos ainda insistem que os finlandeses são "predominantemente do Leste Europeu e compostos por pessoas que migraram para o norte do refúgio glacial na Ucrânia durante a Idade do Gelo".[66] Mais a leste, a questão é menos polêmica. Os portadores do Haplogrupo N representam uma parte significativa de todos os grupos étnicos não-eslavos do norte da Rússia, incluindo 37% dos carélios, 35% dos komis (65% de acordo com outro estudo[67]), 67% dos maris, 98% dos nenets, 94% dos nganasans e 86% a 94% dos iacutes.[68]
As variações genéticas para a tolerância à lactose e maior estatura dos europeus vieram com os proto-indo-europeus.[37]
Durante o Império Romano, as fontes históricas mostram a existência de muitos movimentos migratórios pela Europa, tanto dentro como fora dos limites do Império. As fontes às vezes citam casos de genocídio de tribos de províncias rebeldes atribuídos aos romanos. Se isso de fato ocorreu, teria sido limitado, já que que as populações modernas apresentam considerável continuidade genética em suas respectivas regiões. O processo de romanização aparenta ter sido realizado pela colonização das províncias romanas por um pequeno número de colonos, como administradores, militares, veteranos estabelecidos e comerciantes que emanaram das várias regiões do Império e não apenas da Itália. Eles serviram como um núcleo para a aculturação de notáveis locais.[77] Dado o pequeno número e origens variadas, a romanização parece não ter deixado assinaturas genéticas distintas na Europa. As populações falantes de línguas românicas nos Balcãs, como os romenos, arromenos e moldavos, são geneticamente mais próximas aos seus vizinhos falantes de idiomas helênicos e eslavos do sul, ao invés de italianos modernos.[78][79] Steven Bird especulou que E1b1b1a se espalhou pela Europa durante a época romana, através de populações vindas da Trácia e da Dácia.[57]
Haplogrupo R1a, quase inteiramente no subclado R1a1a, é prevalente em grande parte da Europa Central e Leste Europeu e também no Ásia Meridional e Central. Por exemplo: há um aumento acentuado em R1a1 e uma diminuição em R1b1b2 à medida que se vai para o leste da Alemanha para a Polônia.[88] Também tem uma grande presença na Escandinávia (particularmente na Noruega).[89][90] Nos países bálticos, as frequências R1a diminuem da Lituânia (45%) para a Estônia (cerca de 30%).[91]
E1b1b (anteriormente conhecido como E3b) representa a última grande migração direta da África para a Europa. Acredita-se que apareceu pela primeira vez no Chifre da África há aproximadamente 26 mil anos e se dispersou para o Norte da África e o Oriente Próximo durante os períodos paleolítico e mesolítico tardios. As linhagens E1b1b estão intimamente ligadas à difusão das línguas afro-asiáticas. Embora presente em toda o continente europeu, atinge o pico nos Balcãs ocidentais, entre os albaneses e seus vizinhos. Também é comum na Itália e na Península Ibérica. O haplogrupo E1b1b1, principalmente na forma de seu subclado E1b1b1a2 (E-V13), atinge frequências acima de 47% ao redor da área de Kosovo.[33] Acredita-se que este clado tenha chegado à Europa a partir do Oriente Próximo no Mesolítico posterior,[54] ou no Neolítico.[92] O subclado E-M81 do Norte da África também está presente na Sicília e na Andaluzia.
Ao deixar de lado todos os pequenos enclaves, há também vários haplogrupos além dos quatro acima citados que são mais comuns apenas em certas áreas da Europa.
Haplogrupo G, pertencente aos europeus neolíticos originais, é comum na maior parte da Europa em frequências baixas, atingindo o máximo acima de 70% na região da Geórgia e entre os madjars do Cazaquistão (embora vivam na Ásia, os madjars fazem fronteira com o perímetro oriental da Europa), até 10% na Sardenha, 12% na Córsega e Uppsala, 11% nos Balcãs e Portugal, 10% na Espanha e 9% na Rússia europeia. Este clado também é encontrado no Oriente Próximo.
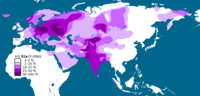
Haplogrupo N é comum somente no nordeste da Europa e, na forma de seu subclado N1c1, atinge frequências de aproximadamente 60% entre os finlandeses e aproximadamente 40% entre os povos bálticos e estonianos.
Haplogrupo J2, em vários subclados (J2a, J2b), é encontrado em níveis de cerca de 15 a 30% nos Balcãs (particularmente na Grécia) e na Itália, sendo também frequente no Oriente Médio.[93]
Distribuição dos haplogrupos de cromossomo Y mais comuns na Europa
Mapa e gráfico em alemão mostrando a frequência de haplogrupos de DNA mitocondrial na Europa e regiões próximas
Houve um grande número de estudos sobre os haplogrupos de DNA mitocondrial na Europa. Em contraste com os haplogrupos do cromossomo Y, os haplogrupos de DNA mitocondrial não mostraram uma padronização de acordo com o local, mas eram mais uniformemente ubíquos. Para além dos periféricos lapões, todos os europeus são caracterizados pela predominância dos haplogrupos H, U e T. A falta de um padrão geográfico observável do DNA mitocondrial provavelmente se deve a fatores socioculturais, como fenómenos de poliginia e patrilocalidade.[51]
Os estudos genéticos sugerem algum fluxo dos genes de DNA mitocondrial para a Europa Oriental vindos da Ásia Oriental ou do Sul da Sibéria há entre 13000 e 6000 anos.[94] A análise de esqueletos neolíticos da Grande Planície Húngara encontrou uma alta frequência de haplogrupos de DNA mitocondrial da Ásia Oriental, alguns dos quais sobrevivem em populações modernas do leste europeu.[94] O fluxo gênico materno para a Europa da África Subsaariana começou há 11 mil anos, embora aproximadamente 65% das linhagens tenham chegado mais recentemente, inclusive durante o período de romanização, as expansões árabes do sul da Europa e durante o Tráfico de escravos no Atlântico.[95]
Seldin (2006) usou mais de 5.000 SNPs autossômicos, mostrando "uma distinção consistente e reproduzível entre as populações europeias do norte e do sul do continente". A maioria dos participantes individuais com ascendência do sul da Europa, como italianos, gregos, portugueses e espanhóis, e judeus asquenazes têm mais de 85% de membros em populações do sul; e a maioria dos europeus do norte, oeste, centro e leste, como suecos, ingleses, irlandeses, alemães e ucranianos, têm mais de 90% no grupo populacional do norte. Muitos dos participantes deste estudo eram cidadãos estadunidenses que se identificaram com diferentes etnias europeias com base no pedigree familiar auto-relatado.[96]
Um estudo semelhante em 2007, o qual utilizou-se de amostras predominantemente vindas da Europa, descobriu que a diferenciação genética mais importante no continente europeu ocorre em uma linha do norte ao sudeste (norte da Europa aos Balcãs), com outro eixo leste-oeste de diferenciação em todo o continente. Suas descobertas foram consistentes com resultados anteriores baseados em DNA mitocondrial e DNA do cromossomo Y que apoiam a teoria de que as populações nativas modernas da Península Ibérica possuem a ancestralidade genética europeia mais antiga, além de separar bascos e lapões de outras populações europeias.[97]
Foi sugerido que os ingleses e irlandeses fossem agrupados com outras etnias europeias do norte e do leste, como alemães e poloneses, enquanto alguns indivíduos bascos e italianos também são agrupados com os europeus do norte. Apesar dessas estratificações, foi observado que "há baixa diversidade aparente na Europa, com amostras de todo o continente apenas marginalmente mais dispersas do que amostras de populações únicas em outras partes do mundo".[97]
Em 2008, duas equipes internacionais de pesquisa publicaram análises dos genótipos de muitos europeus, utilizando mais de 300.000 SNPs autossômicos. Com exceção de grupos étnicos isolados já conhecidos, como os bascos, finlandeses e sardos, a população europeia não possuía grandes descontinuidades acentuadas como encontradas em estudos anteriores (por exemplo: Seldin et al., 2006 e Bauchet et al., 2007[97]), apesar de existir uma gradiente distinguível de sul a norte. No geral, encontraram apenas baixa diferenciação genética entre as subpopulações, e as diferenças existentes foram caracterizadas por uma forte correlação continental entre a distância geográfica e genética. Além disso, eles descobriram que a diversidade era maior no sul da Europa, devido a um tamanho populacional efetivo maior e/ou expansão populacional do sul para o norte da Europa.[98] Os pesquisadores entendem que essa observação implica que, geneticamente, os europeus não são distribuídos em populações distintas.[99][98]
Dois estudos de genoma completo de ucranianos e russos mostraram que a diversidade genômica desta região não foi representada nas pesquisas genéticas anteriores, visto que os estudos gênicos na Europa são mais tendenciosos para populações da parte ocidental do continente.[100][101] No nordeste da Rússia, os komis, falantes de um idioma das línguas urálicas, formam um polo de diversidade genética difrente de outras populações, deslocado para populações relacionadas à Ásia Oriental.[102][69]
De acordo com o geneticista David Reich, com base em genomas humanos antigos sequenciados por seu laboratório em 2016, os europeus são fruto da mistura de quatro componentes ancestrais: WHG (caçadores-coletores ocidentais), EHG (Caçadores-Coletores orientais) e EEF (primeiros agricultores europeus, vindos da Anatólia) em graus variados.[103] Foi descoberto também que os caçadores-coletores orientais também carregavam alguma ascendência asiática.[104]
↑ abSeguin-Orlando A, Korneliussen TS, Sikora M, Malaspinas AS, Manica A, Moltke I, et al. (novembro de 2014). «Paleogenomics. Genomic structure in Europeans dating back at least 36,200 years». Science (em inglês). 346 (6213): 1113–1118. Bibcode:2014Sci...346.1113S. PMID25378462. doi:10.1126/science.aaa0114
↑Posth C, Renaud G, Mittnik A, Drucker DG, Rougier H, Cupillard C, et al. (junho de 2016). «Pleistocene Mitochondrial Genomes Suggest a Single Major Dispersal of Non-Africans and a Late Glacial Population Turnover in Europe». Current Biology (em inglês). 26 (6): 827–833. PMID26853362. doi:10.1016/j.cub.2016.01.037
↑ abDesde Lazaridis et al. (2014), outros estudos evidenciaram o quadro de mistura entre EEF e WHG. Em uma análise de 2017 de 180 genomas antigos dos períodos Calcolítico e Neolítico da Hungria, Alemanha e Península Ibérica, foram encontradas evidências de um período prolongado de cruzamento EEF-WHG. A miscigenação ocorreu regionalmente, a partir de populações locais de caçadores-coletores, de modo que as populações das três regiões citadas eram geneticamente distinguíveis em todas as fases do Neolítico, com uma proporção gradualmente crescente de ascendência WHG de populações agrícolas ao longo do tempo. Isso sugere que, após a expansão inicial dos primeiros agricultores vindos do Oriente Próximo, não houve mais migrações de longo alcance substanciais o suficiente para homogeneizar a população agrícola, e que as populações agrícolas e de caçadores-coletores existiram lado a lado por muitos séculos, com uma mistura gradual e contínua ao longo do Quinto e Quarto milênios antes de Cristo, ao invés de um único evento de mistura no contato inicial.
Lipson M, Szécsényi-Nagy A, Mallick S, Pósa A, Stégmár B, Keerl V, et al. (novembro de 2017). «Parallel palaeogenomic transects reveal complex genetic history of early European farmers». Nature (em inglês). 551 (7680): 368–372. Bibcode:2017Natur.551..368L. PMC5973800. PMID29144465. doi:10.1038/nature24476
↑Mesmo antes do aparecimento dos estudos genéticos, alguns antropólogos acreditavam ter descoberto esqueletos híbridos entre humanos modernos e neandertais. Esses resultados foram considerados "ambíguos". Evidências arqueológicas apontam para uma mudança abrupta de artefatos neandertais para aqueles relacionados aos humanos anatomicamente modernos durante o Paleolítico Superior. Klein RG (março de 2003). «Paleoanthropology. Whither the Neanderthals?». Science. 299 (5612): 1525–1527. PMID12624250. doi:10.1126/science.1082025
↑«Homo neanderthalensis». The Smithsonian Institution's Human Origins Program (em inglês). Consultado em 15 de dezembro de 2023
↑Hardy J, Pittman A, Myers A, Gwinn-Hardy K, Fung HC, de Silva R, et al. (agosto de 2005). «Evidence suggesting that Homo neanderthalensis contributed the H2 MAPT haplotype to Homo sapiens». Biochemical Society Transactions (em inglês). 33 (Pt 4): 582–585. PMID16042549. doi:10.1042/bst0330582
↑Shaw-Smith C, Pittman AM, Willatt L, Martin H, Rickman L, Gribble S, et al. (setembro de 2006). «Microdeletion encompassing MAPT at chromosome 17q21.3 is associated with developmental delay and learning disability». Nature Genetics (em inglês). 38 (9): 1032–1037. PMID16906163. doi:10.1038/ng1858
↑Almos PZ, Horváth S, Czibula A, Raskó I, Sipos B, Bihari P, et al. (novembro de 2008). «H1 tau haplotype-related genomic variation at 17q21.3 as an Asian heritage of the European Gypsy population». Heredity (em inglês). 101 (5): 416–419. PMID18648385. doi:10.1038/hdy.2008.70
↑ abEuropean Prehistory: a survey (em inglês). [S.l.]: Birkhauser. 2002. ISBN978-0-306-46793-6
↑ abcdefgSemino O, Passarino G, Oefner PJ, Lin AA, Arbuzova S, Beckman LE, et al. (novembro de 2000). «The genetic legacy of Paleolithic Homo sapiens sapiens in extant Europeans: a Y chromosome perspective». Science (em inglês). 290 (5494): 1155–1159. Bibcode:2000Sci...290.1155S. PMID11073453. doi:10.1126/science.290.5494.1155 Observação: Os nomes dos haplogrupos estão grafados diferentes neste artigo científico. Por exemplo: o haplogrupo I é referido como "M170".
↑ abcPericić M, Lauc LB, Klarić IM, Rootsi S, Janićijevic B, Rudan I, et al. (outubro de 2005). «High-resolution phylogenetic analysis of southeastern Europe traces major episodes of paternal gene flow among Slavic populations». Molecular Biology and Evolution (em inglês). 22 (10): 1964–1975. PMID15944443. doi:10.1093/molbev/msi185
↑Runnel, C. (2003). “The Origins of the Greek Neolithic: A Personal View”. In: Ammerman, A. J. e Biagi, P. (eds.): The Widening Harvest. The Neolithic Transition in Europe: Looking Back, Looking Forward. Archaeological Institute of America, Boston, 121-132.
↑Underhill PA, Kivisild T (2007). «Use of y chromosome and mitochondrial DNA population structure in tracing human migrations». Annual Review of Genetics (em inglês). 41: 539–564. PMID18076332. doi:10.1146/annurev.genet.41.110306.130407
↑Y chromosome data show a signal for a separate late-Pleistocene migration from Africa to Europe via Sinai as evidenced through the distribution of haplogroup E3b lineages, which is not manifested in mtDNA haplogroup distributions.
↑Allentoft, Morten E.; Sikora, Martin; Sjögren, Karl-Göran; Rasmussen, Simon; Rasmussen, Morten; Stenderup, Jesper; Damgaard, Peter B.; Schroeder, Hannes; Ahlström, Torbjörn (junho de 2015). «Population genomics of Bronze Age Eurasia». Nature (em inglês). 522 (7555): 167–172. ISSN1476-4687. doi:10.1038/nature14507. Consultado em 7 de julho de 2023
↑Ingman M, Gyllensten U (janeiro de 2007). «A recent genetic link between Sami and the Volga-Ural region of Russia». European Journal of Human Genetics (em inglês). 15 (1): 115–120. PMID16985502. doi:10.1038/sj.ejhg.5201712
↑«Who Are the Finns?». A Man of Measure Festschrift in Honour of Fred Karlsson (em inglês). [S.l.: s.n.] pp. 97–108|acessodata= requer |url= (ajuda)
↑ abMirabal S, Varljen T, Gayden T, Regueiro M, Vujovic S, Popovic D, et al. (julho de 2010). «Human Y-chromosome short tandem repeats: a tale of acculturation and migrations as mechanisms for the diffusion of agriculture in the Balkan Peninsula». American Journal of Physical Anthropology (em inglês). 142 (3): 380–390. PMID20091845. doi:10.1002/ajpa.21235
↑Rootsi S, Zhivotovsky LA, Baldovic M, Kayser M, Kutuev IA, Khusainova R, et al. (fevereiro de 2007). «A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe». European Journal of Human Genetics (em inglês). 15 (2): 204–211. PMID17149388. doi:10.1038/sj.ejhg.5201748, Tabela complementar 1.
↑Pannonia and Upper Moesia: a history of the middle Danube provinces of the Roman Empire (em inglês). London: Routledge & Kegan Paul. 1974. ISBN978-0-7100-7714-1
↑Comas D, Schmid H, Braeuer S, Flaiz C, Busquets A, Calafell F, et al. (março de 2004). «Alu insertion polymorphisms in the Balkans and the origins of the Aromuns». Annals of Human Genetics (em inglês). 68 (Pt 2): 120–127. PMID15008791. doi:10.1046/j.1529-8817.2003.00080.x
↑Bosch E, Calafell F, González-Neira A, Flaiz C, Mateu E, Scheil HG, et al. (julho de 2006). «Paternal and maternal lineages in the Balkans show a homogeneous landscape over linguistic barriers, except for the isolated Aromuns». Annals of Human Genetics. 70 (Pt 4): 459–487. PMID16759179. doi:10.1111/j.1469-1809.2005.00251.x
↑Kasperaviciūte D, Kucinskas V, Stoneking M (setembro de 2004). «Y chromosome and mitochondrial DNA variation in Lithuanians». Annals of Human Genetics (em inglês). 68 (Pt 5): 438–52. PMID15469421. doi:10.1046/j.1529-8817.2003.00119.x
↑ abLao O, Lu TT, Nothnagel M, Junge O, Freitag-Wolf S, Caliebe A, et al. (agosto de 2008). «Correlation between genetic and geographic structure in Europe». Current Biology (em inglês). 18 (16): 1241–1248. PMID18691889. doi:10.1016/j.cub.2008.07.049






,_The_Horse,_The_Wheel_and_Language.jpg)





.png)


 . PMID 30837540. doi:10.1038/s41598-019-39802-1
. PMID 30837540. doi:10.1038/s41598-019-39802-1