
Córtex cerebral
| Cortex Cerebral | |||
|---|---|---|---|
| Cortex cerebri | |||
|
Partes do Encéfalo | |||
 | |||
| Localização | Encefalo | ||
| Sistema | Sistema nervoso central | ||
| Veia | Veia cerebral magna | ||
| Artéria | Artéria cerebral anterior, Artéria cerebral média e Artéria cerebral posterior | ||
| Componentes | Lobo frontal, Lobo parietal, Lobo temporal, Lobo occipital, Ínsula e Lobo límbico | ||
| Função | Pensamento, Movimento voluntário, Linguagem, Julgamento e Percepção | ||
| Latim | Cortex cerebri | ||
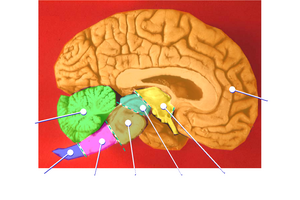
O córtex cerebral, também conhecido como manto cerebral,[1] é a camada externa do tecido neural do cérebro em humanos e outros mamíferos . O córtex cerebral consiste principalmente no neocórtex de seis camadas, com apenas 10% consistindo no alocórtex .[2] É separado em dois córtices, pela fissura longitudinal que divide o cérebro nos hemisférios cerebrais esquerdo e direito. Os dois hemisférios são unidos abaixo do córtex pelo corpo caloso . O córtex cerebral é o maior local de integração neural no sistema nervoso central .[3] Desempenha um papel fundamental na atenção, percepção, consciência, pensamento, memória, linguagem e consciência. O córtex cerebral é a parte do cérebro responsável pela cognição.
Na maioria dos mamíferos, com exceção dos pequenos mamíferos que possuem cérebros pequenos, o córtex cerebral é dobrado, proporcionando uma maior área de superfície no volume confinado do crânio . Além de minimizar o volume cerebral e craniano, o dobramento cortical é crucial para os circuitos cerebrais e sua organização funcional.[4] Nos mamíferos com cérebros pequenos não há dobramento e o córtex é liso.[5][6]
Uma dobra ou crista no córtex é denominada giro (plural giros) e um sulco é denominado sulco (plural sulcos). Estas circunvoluções superficiais aparecem durante o desenvolvimento fetal e continuam a amadurecer após o nascimento através do processo de girificação . No cérebro humano, a maior parte do córtex cerebral não é visível do exterior, mas está enterrado nos sulcos.[7] Os principais sulcos e giros marcam as divisões do cérebro nos lobos do cérebro . Os quatro lobos principais são os lobos frontal, parietal, occipital e temporal . Outros lobos são o lobo límbico e o córtex insular, muitas vezes referido como lobo insular.
Existem entre 14 e 16 bilhões de neurônios no córtex cerebral humano.[8] Estas são organizadas em camadas corticais horizontais, e radialmente em colunas e minicolunas corticais . As áreas corticais têm funções específicas, como movimento no córtex motor e visão no córtex visual . O córtex motor está localizado principalmente no giro pré-central, e o córtex visual está localizado no lobo occipital.
Estrutura
[editar | editar código-fonte]
O córtex cerebral é a cobertura externa das superfícies dos hemisférios cerebrais e é dobrado em picos chamados giros e sulcos chamados sulcos . No cérebro humano está entre 2 e 3-4 mm. grosso,[9] e representa 40% da massa do cérebro.[10] 90% do córtex cerebral é o neocórtex de seis camadas, enquanto os outros 10% são compostos de alocórtex de três/quatro camadas.[10] Existem entre 14 e 16 bilhões de neurônios no córtex,[10] e estes estão organizados radialmente em colunas corticais, e minicolunas, nas camadas organizadas horizontalmente do córtex.[11][12]
O neocórtex é separável em diferentes regiões do córtex conhecidas no plural como córtices, e incluem o córtex motor e o córtex visual . Cerca de dois terços da superfície cortical estão enterrados nos sulcos e o córtex insular está completamente oculto. O córtex é mais espesso na parte superior do giro e mais fino na parte inferior do sulco.[13]
Dobras anatômicas
[editar | editar código-fonte]O córtex cerebral é dobrado de uma forma que permite que uma grande área superficial de tecido neural caiba dentro dos limites do neurocrânio . Quando desdobrado no ser humano, cada córtex hemisférico tem uma área de superfície total de cerca de 0,12 metro quadrados (1,3 sq ft) .[14] A dobra é voltada para dentro, longe da superfície do cérebro, e também está presente na superfície medial de cada hemisfério dentro da fissura longitudinal . A maioria dos mamíferos tem um córtex cerebral que é enrolado com picos conhecidos como giros e depressões ou sulcos conhecidos como sulcos. Alguns pequenos mamíferos, incluindo alguns pequenos roedores, têm superfícies cerebrais lisas sem girificação.[15]
Lobos
[editar | editar código-fonte]Os sulcos e giros maiores marcam as divisões do córtex do cérebro nos lobos do cérebro .[16] Existem quatro lobos principais: o lobo frontal, o lobo parietal, o lobo temporal e o lobo occipital . O córtex insular é frequentemente incluído como lobo insular.[17] O lobo límbico é uma borda do córtex no lado medial de cada hemisfério e também é frequentemente incluído.[18] Existem também três lóbulos cerebrais descritos: o lóbulo paracentral, o lóbulo parietal superior e o lóbulo parietal inferior .
Grossura
[editar | editar código-fonte]Para espécies de mamíferos, cérebros maiores (em termos absolutos, não apenas em relação ao tamanho corporal) tendem a ter córtices mais espessos.[19] Os menores mamíferos, como os musaranhos, têm uma espessura neocortical de cerca de 0,5 milímetros; aqueles com os maiores cérebros, como humanos e baleias-comuns, têm espessuras de 2–4 milímetros.[20][21] Existe uma relação aproximadamente logarítmica entre o peso do cérebro e a espessura cortical.[19] A ressonância magnética do cérebro (MRI) permite medir a espessura do córtex cerebral humano e relacioná-la com outras medidas. A espessura das diferentes áreas corticais varia, mas em geral o córtex sensorial é mais fino que o córtex motor.[22] Um estudo encontrou alguma associação positiva entre a espessura cortical e a inteligência.[23] Outro estudo descobriu que o córtex somatossensorial é mais espesso em pacientes com enxaqueca, embora não se saiba se isso é resultado de crises de enxaqueca, a causa delas ou se ambas são resultado de uma causa compartilhada.[24][25] Um estudo posterior utilizando uma população maior de pacientes não relatou nenhuma alteração na espessura cortical em pacientes com enxaqueca.[26] Um distúrbio genético do córtex cerebral, em que a diminuição do dobramento em certas áreas resulta em um microgiro, onde há quatro camadas em vez de seis, é em alguns casos visto como relacionado à dislexia .[27]
Camadas de neocórtex
[editar | editar código-fonte]
O neocórtex é formado por seis camadas, numeradas de I a VI, desde a camada mais externa I - próxima à pia-máter, até a camada mais interna VI - próxima à substância branca subjacente. Cada camada cortical possui uma distribuição característica de diferentes neurônios e suas conexões com outras regiões corticais e subcorticais. Existem conexões diretas entre diferentes áreas corticais e conexões indiretas através do tálamo.
Um dos exemplos mais claros de estratificação cortical é a linha de Gennari no córtex visual primário . É uma faixa de tecido mais branco que pode ser observada a olho nu no sulco calcarino do lobo occipital. A linha de Gennari é composta por axônios que trazem informações visuais do tálamo para a camada IV do córtex visual .
A coloração de seções transversais do córtex para revelar a posição dos corpos celulares neuronais e dos tratos axônicos intracorticais permitiu que os neuroanatomistas no início do século 20 produzissem uma descrição detalhada da estrutura laminar do córtex em diferentes espécies. O trabalho de Korbinian Brodmann (1909) estabeleceu que o neocórtex dos mamíferos é consistentemente dividido em seis camadas.
Primeira camada
[editar | editar código-fonte]A camada I é a camada molecular, e contém poucos neurônios dispersos, incluindo neurônios GABAérgicos de rosa mosqueta.[28] A camada I consiste em grande parte em extensões de tufos dendríticos apicais de neurônios piramidais e axônios orientados horizontalmente, bem como células gliais .[29] Durante o desenvolvimento, células de Cajal-Retzius[30] e células da camada granular subpial[31] estão presentes nesta camada. Além disso, algumas células estreladas espinhosas podem ser encontradas aqui. Acredita-se que as entradas para os tufos apicais sejam cruciais para as interações de feedback no córtex cerebral envolvidas na aprendizagem associativa e na atenção.[32] Embora se pensasse que a entrada para a camada I vinha do próprio córtex,[33] agora percebe-se que a camada I através do manto do córtex cerebral recebe informações substanciais da matriz ou das células do tálamo do tipo M[34] (em contraste com core ou tipo C que vão para a camada IV).[35]
Segunda camada
[editar | editar código-fonte]A camada II, a camada granular externa, contém pequenos neurônios piramidais e numerosos neurônios estrelados.
Terceira camada
[editar | editar código-fonte]A camada III, a camada piramidal externa, contém predominantemente neurônios piramidais de tamanho pequeno e médio, bem como neurônios não piramidais com axônios intracorticais orientados verticalmente; as camadas I a III são o principal alvo dos aferentes corticocorticais inter-hemisféricos, e a camada III é a principal fonte dos eferentes corticocorticais.
Quarta camada
[editar | editar código-fonte]A camada IV, a camada granular interna, contém diferentes tipos de células estreladas e piramidais e é o principal alvo dos aferentes tálamo-corticais dos neurônios tálamo tipo C (tipo núcleo),[36] bem como dos aferentes corticocorticais intra-hemisféricos. As camadas acima da camada IV também são chamadas de camadas supragranulares (camadas I-III), enquanto as camadas abaixo são chamadas de camadas infragranulares (camadas V e VI).
Quinta camada
[editar | editar código-fonte]A camada V, a camada piramidal interna, contém grandes neurônios piramidais. Os axônios destes deixam o córtex e se conectam com estruturas subcorticais, incluindo os gânglios da base . No córtex motor primário do lobo frontal, a camada V contém células piramidais gigantes chamadas células de Betz, cujos axônios viajam através da cápsula interna, do tronco cerebral e da medula espinhal formando o trato corticoespinhal, que é a principal via para o controle motor voluntário.
Sexta camada
[editar | editar código-fonte]A camada VI, a camada polimórfica ou multiforme, contém poucos neurônios piramidais grandes e muitos neurônios piramidais e multiformes pequenos, fusiformes; a camada VI envia fibras eferentes ao tálamo, estabelecendo uma interconexão recíproca muito precisa entre o córtex e o tálamo.[37] Ou seja, os neurônios da camada VI de uma coluna cortical conectam-se aos neurônios do tálamo que fornecem entrada para a mesma coluna cortical. Essas conexões são excitatórias e inibitórias. Os neurônios enviam fibras excitatórias aos neurônios do tálamo e também enviam colaterais ao núcleo reticular do tálamo que inibem esses mesmos neurônios do tálamo ou adjacentes a eles.[38] Uma teoria é que, como a saída inibitória é reduzida pela entrada colinérgica no córtex cerebral, isso fornece ao tronco cerebral um "controle de ganho ajustável para a retransmissão de entradas lemniscais ".[38]
Fornecimento de sangue e drenagem
[editar | editar código-fonte]
O suprimento sanguíneo para o córtex cerebral faz parte da circulação cerebral . As artérias cerebrais fornecem o sangue que perfunde o cérebro. Este sangue arterial transporta oxigênio, glicose e outros nutrientes para o córtex. As veias cerebrais drenam o sangue desoxigenado e os resíduos metabólicos, incluindo o dióxido de carbono, de volta ao coração.
As principais artérias que irrigam o córtex são a artéria cerebral anterior, a artéria cerebral média e a artéria cerebral posterior . A artéria cerebral anterior irriga as porções anteriores do cérebro, incluindo a maior parte do lobo frontal. A artéria cerebral média supre os lobos parietais, lobos temporais e partes dos lobos occipitais. A artéria cerebral média se divide em dois ramos para suprir os hemisférios esquerdo e direito, onde se ramificam ainda mais. A artéria cerebral posterior irriga os lobos occipitais. O círculo de Willis é o principal sistema sanguíneo que lida com o suprimento de sangue no cérebro e no córtex cerebral.
Desenvolvimento
[editar | editar código-fonte]O desenvolvimento pré-natal do córtex cerebral é um processo complexo e afinado denominado corticogênese, influenciado pela interação entre os genes e o meio ambiente.[39]
Tubo neural
[editar | editar código-fonte]O córtex cerebral se desenvolve a partir da parte mais anterior, a região do prosencéfalo, do tubo neural .[40][41] A placa neural dobra-se e fecha-se para formar o tubo neural . A partir da cavidade dentro do tubo neural desenvolve-se o sistema ventricular e, a partir das células neuroepiteliais de suas paredes, os neurônios e glia do sistema nervoso. A parte mais anterior (frontal ou cranial) da placa neural, o prosencéfalo, que é evidente antes do início da neurulação, dá origem aos hemisférios cerebrais e, posteriormente, ao córtex.[42]
Desenvolvimento de neurônios corticais
[editar | editar código-fonte]Os neurônios corticais são gerados dentro da zona ventricular, próximo aos ventrículos . A princípio, esta zona contém células-tronco neurais, que transitam para células gliais radiais – células progenitoras, que se dividem para produzir células gliais e neurônios.[43]
Gália radial
[editar | editar código-fonte]O córtex cerebral é composto por uma população heterogênea de células que dão origem a diferentes tipos de células. A maioria dessas células é derivada da migração radial da glia que forma os diferentes tipos de células do neocórtex e é um período associado ao aumento da neurogênese . Da mesma forma, o processo de neurogênese regula a laminação para formar as diferentes camadas do córtex. Durante este processo há um aumento na restrição do destino celular que começa com os progenitores anteriores dando origem a qualquer tipo de célula no córtex e os progenitores posteriores dando origem apenas aos neurônios das camadas superficiais. Esse destino celular diferencial cria uma topografia de dentro para fora no córtex, com neurônios mais jovens nas camadas superficiais e neurônios mais velhos nas camadas mais profundas. Além disso, os neurônios laminares são parados na fase S ou G2 para fornecer uma distinção precisa entre as diferentes camadas corticais. A diferenciação laminar não está totalmente completa até depois do nascimento, uma vez que durante o desenvolvimento os neurônios laminares ainda são sensíveis a sinais extrínsecos e estímulos ambientais.[44]
Evolução
[editar | editar código-fonte]De todas as diferentes regiões cerebrais, o córtex cerebral apresenta a maior variação evolutiva e evoluiu mais recentemente.[45] Em contraste com o circuito altamente conservado da medula oblonga, por exemplo, que desempenha funções críticas como a regulação das frequências cardíaca e respiratória, muitas áreas do córtex cerebral não são estritamente necessárias para a sobrevivência. Assim, a evolução do córtex cerebral viu o advento e a modificação de novas áreas funcionais – particularmente áreas de associação que não recebem informações diretamente de fora do córtex.[45]
Uma teoria chave da evolução cortical está incorporada na hipótese da unidade radial e na hipótese do protomapa relacionada, proposta pela primeira vez por Rakic.[46] Esta teoria afirma que novas áreas corticais são formadas pela adição de novas unidades radiais, o que é realizado ao nível das células estaminais . A hipótese do protomapa afirma que a identidade celular e molecular e as características dos neurônios em cada área cortical são especificadas por células-tronco corticais, conhecidas como células gliais radiais, em um mapa primordial. Este mapa é controlado por proteínas de sinalização secretadas e fatores de transcrição a jusante.[47][48][49]
Função
[editar | editar código-fonte]
Conexões
[editar | editar código-fonte]O córtex cerebral está conectado a diversas estruturas subcorticais, como o tálamo e os gânglios da base, enviando-lhes informações através de conexões eferentes e recebendo deles informações por meio de conexões aferentes . A maior parte da informação sensorial é encaminhada para o córtex cerebral através do tálamo. A informação olfativa, entretanto, passa através do bulbo olfatório até o córtex olfatório ( córtex piriforme ). A maioria das conexões ocorre de uma área do córtex para outra, e não de áreas subcorticais; Braitenberg e Schüz (1998) afirmam que em áreas sensoriais primárias, no nível cortical onde terminam as fibras de entrada, até 20% das sinapses são fornecidas por aferentes extracorticais, mas que em outras áreas e outras camadas a porcentagem é provavelmente muito maior. mais baixo.[50]
Áreas corticais
[editar | editar código-fonte]Todo o córtex cerebral foi dividido em 52 áreas diferentes numa apresentação inicial de Korbinian Brodmann . Estas áreas, conhecidas como áreas de Brodmann, baseiam-se na sua citoarquitetura, mas também se relacionam com diversas funções. Um exemplo é a área 17 de Brodmann, que é o córtex visual primário .
Em termos mais gerais, o córtex é normalmente descrito como compreendendo três partes: áreas sensoriais, motoras e de associação.
Áreas sensoriais
[editar | editar código-fonte]As áreas sensoriais são as áreas corticais que recebem e processam informações dos sentidos . Partes do córtex que recebem informações sensoriais do tálamo são chamadas de áreas sensoriais primárias. Os sentidos da visão, audição e tato são atendidos pelo córtex visual primário, córtex auditivo primário e córtex somatossensorial primário, respectivamente. Em geral, os dois hemisférios recebem informações do lado oposto (contralateral) do corpo . Por exemplo, o córtex somatossensorial primário direito recebe informações dos membros esquerdos, e o córtex visual direito recebe informações do campo visual esquerdo. A organização dos mapas sensoriais no córtex reflete a do órgão sensorial correspondente, no que é conhecido como mapa topográfico . Os pontos vizinhos no córtex visual primário, por exemplo, correspondem a pontos vizinhos na retina . Este mapa topográfico é denominado mapa retinotópico . Da mesma forma, existe um mapa tonotópico no córtex auditivo primário e um mapa somatotópico no córtex sensorial primário. Este último mapa topográfico do corpo no giro central posterior foi ilustrado como uma representação humana deformada, o homúnculo somatossensorial, onde o tamanho das diferentes partes do corpo reflete a densidade relativa de sua inervação. Áreas com muita inervação sensorial, como as pontas dos dedos e os lábios, requerem mais área cortical para processar sensações mais refinadas.
Áreas motoras
[editar | editar código-fonte]As áreas motoras estão localizadas em ambos os hemisférios do córtex. As áreas motoras estão intimamente relacionadas com o controle dos movimentos voluntários, especialmente os movimentos finos e fragmentados executados pela mão. A metade direita da área motora controla o lado esquerdo do corpo e vice-versa.
Logo abaixo do córtex cerebral estão massas subcorticais interconectadas de substância cinzenta chamadas gânglios da base (ou núcleos). Os gânglios da base recebem informações da substância negra do mesencéfalo e das áreas motoras do córtex cerebral e enviam sinais de volta para ambos os locais. Eles estão envolvidos no controle motor. Eles são encontrados lateralmente ao tálamo. Os principais componentes dos gânglios da base são o núcleo caudado, o putâmen, o globo pálido, a substância negra, o núcleo accumbens e o núcleo subtalâmico . O putâmen e o globo pálido também são conhecidos coletivamente como núcleo lentiforme, porque juntos formam um corpo em forma de lente. O putâmen e o núcleo caudado também são chamados coletivamente de corpo estriado devido à sua aparência listrada.[51][52]
Áreas de associação
[editar | editar código-fonte]As áreas de associação são as partes do córtex cerebral que não pertencem às regiões primárias. Eles funcionam para produzir uma experiência perceptual significativa do mundo, permitem-nos interagir de forma eficaz e apoiam o pensamento e a linguagem abstratos. Os lobos parietal, temporal e occipital – todos localizados na parte posterior do córtex – integram informações sensoriais e informações armazenadas na memória. O lobo frontal ou complexo de associação pré-frontal está envolvido no planejamento de ações e movimentos, bem como no pensamento abstrato. Globalmente, as áreas de associação estão organizadas como redes distribuídas.[53] Cada rede conecta áreas distribuídas em regiões amplamente espaçadas do córtex. Redes distintas são posicionadas adjacentes umas às outras, produzindo uma série complexa de redes entrelaçadas. A organização específica das redes associativas é debatida com evidências de interações, relações hierárquicas e competição entre redes.
Nos humanos, as redes de associação são particularmente importantes para o funcionamento da linguagem. No passado, foi teorizado que as habilidades linguísticas estão localizadas na área de Broca, em áreas do giro frontal inferior esquerdo, BA44 e BA45, para expressão da linguagem e na área de Wernicke BA22, para recepção da linguagem. No entanto, foi demonstrado que os processos de expressão e recepção da linguagem ocorrem em outras áreas além das estruturas ao redor do sulco lateral, incluindo o lobo frontal, gânglios da base, cerebelo e ponte .[54]
Significado clínico
[editar | editar código-fonte]
Doenças neurodegenerativas, como a doença de Alzheimer, apresentam como marcador uma atrofia da substância cinzenta do córtex cerebral.[56]
Outras doenças do sistema nervoso central incluem distúrbios neurológicos como epilepsia, distúrbios do movimento e diferentes tipos de afasia (dificuldades na expressão ou compreensão da fala).
Danos cerebrais causados por doenças ou traumas podem envolver danos a um lobo específico, como no distúrbio do lobo frontal, e funções associadas serão afetadas. A barreira hematoencefálica que serve para proteger o cérebro contra infecções pode ficar comprometida, permitindo a entrada de patógenos .
O feto em desenvolvimento é suscetível a uma série de fatores ambientais que podem causar defeitos congênitos e problemas no desenvolvimento posterior. O consumo materno de álcool, por exemplo, pode causar distúrbios do espectro alcoólico fetal .[57] Outros fatores que podem causar distúrbios do neurodesenvolvimento são substâncias tóxicas, como medicamentos, e a exposição à radiação, como os raios X. As infecções também podem afetar o desenvolvimento do córtex. Uma infecção viral é uma das causas da lisencefalia, que resulta em um córtex liso sem girificação .
Um tipo de eletrocorticografia denominado mapeamento de estimulação cortical é um procedimento invasivo que envolve a colocação de eletrodos diretamente no cérebro exposto para localizar as funções de áreas específicas do córtex. É usado em aplicações clínicas e terapêuticas, incluindo mapeamento pré-cirúrgico.[58]
Referências
- ↑ «cerebral mantle». TheFreeDictionary.com
- ↑ Strominger, Norman L.; Demarest, Robert J.; Laemle, Lois B. (2012). «Cerebral Cortex». Noback's Human Nervous System, Seventh Edition (em inglês). [S.l.]: Humana Press. pp. 429–451. ISBN 978-1-61779-778-1. doi:10.1007/978-1-61779-779-8_25
- ↑ Saladin, Kenneth (2011). Human anatomy 3rd ed. [S.l.]: McGraw-Hill. pp. 416–422. ISBN 9780071222075
- ↑ Shipp, Stewart (17 de junho de 2007). «Structure and function of the cerebral cortex». Current Biology. 17 (12): R443–9. PMC 1870400
 . PMID 17580069. doi:10.1016/j.cub.2007.03.044
. PMID 17580069. doi:10.1016/j.cub.2007.03.044
- ↑ Fernández, V; Llinares-Benadero, C; Borrell, V (17 de maio de 2016). «Cerebral cortex expansion and folding: what have we learned?». The EMBO Journal. 35 (10): 1021–44. PMC 4868950. PMID 27056680. doi:10.15252/embj.201593701
- ↑ Rakic, P (outubro de 2009). «Evolution of the neocortex: a perspective from developmental biology». Nature Reviews Neuroscience. 10 (10): 724–35. PMC 2913577. PMID 19763105. doi:10.1038/nrn2719
- ↑ Principles of neural science 4th ed. [S.l.]: McGraw-Hill, Health Professions Division. 5 de janeiro de 2000. ISBN 978-0838577011 Verifique o valor de
|url-access=registration(ajuda) - ↑ Saladin, Kenneth (2011). Human anatomy 3rd ed. [S.l.]: McGraw-Hill. pp. 416–422. ISBN 9780071222075
- ↑ Roberts, P (1992). Neuroanatomy 3rd ed. [S.l.]: Springer-Verlag. pp. 86–92. ISBN 9780387977775
- ↑ a b c Saladin, Kenneth (2011). Human anatomy 3rd ed. [S.l.]: McGraw-Hill. pp. 416–422. ISBN 9780071222075
- ↑ Lodato, Simona; Arlotta, Paola (13 de novembro de 2015). «Generating Neuronal Diversity in the Mammalian Cerebral Cortex». Annual Review of Cell and Developmental Biology (em inglês). 31 (1): 699–720. PMC 4778709. PMID 26359774. doi:10.1146/annurev-cellbio-100814-125353
- ↑ Ansen-Wilson, LJ; Lipinski, RJ (janeiro de 2017). «Gene-environment interactions in cortical interneuron development and dysfunction: A review of preclinical studies.». Neurotoxicology. 58: 120–129. PMC 5328258. PMID 27932026. doi:10.1016/j.neuro.2016.12.002
- ↑ Carpenter (1985). Core text of neuroanatomy 3rd ed. [S.l.]: Williams & Wilkins. pp. 348–358. ISBN 978-0683014556
- ↑ Toro, Roberto; Perron, Michel; Pike, Bruce; Richer, Louis; Veillette, Suzanne; Pausova, Zdenka; Paus, Tomáš (1 de outubro de 2008). «Brain Size and Folding of the Human Cerebral Cortex». Cerebral Cortex (em inglês). 18 (10): 2352–2357. ISSN 1047-3211. PMID 18267953. doi:10.1093/cercor/bhm261
- ↑ Rakic, P (outubro de 2009). «Evolution of the neocortex: a perspective from developmental biology». Nature Reviews Neuroscience. 10 (10): 724–35. PMC 2913577. PMID 19763105. doi:10.1038/nrn2719
- ↑ Roberts, P (1992). Neuroanatomy 3rd ed. [S.l.]: Springer-Verlag. pp. 86–92. ISBN 9780387977775
- ↑ Nieuwenhuys, R (2012). The insular cortex: a review. Col: Progress in Brain Research. 195. [S.l.: s.n.] pp. 123–63. PMID 22230626. doi:10.1016/B978-0-444-53860-4.00007-6
- ↑ Tortora, G; Derrickson, B (2011). Principles of anatomy & physiology. 13th. ed. [S.l.]: Wiley. ISBN 9780470646083
- ↑ a b The central nervous system of vertebrates, Volume 1. [S.l.]: Springer. 1998. pp. 2011–2012. ISBN 978-3-540-56013-5
- ↑ Saladin, Kenneth (2011). Human anatomy 3rd ed. [S.l.]: McGraw-Hill. pp. 416–422. ISBN 9780071222075
- ↑ Roberts, P (1992). Neuroanatomy 3rd ed. [S.l.]: Springer-Verlag. pp. 86–92. ISBN 9780387977775
- ↑ Frithjof Kruggel; Martina K. Brückner; Thomas Arendt; Christopher J. Wiggins; D. Yves von Cramon (2003). «Analyzing the neocortical fine-structure». Medical Image Analysis. 7 (3): 251–264. PMID 12946467. doi:10.1016/S1361-8415(03)00006-9
|hdl-access=requer|hdl=(ajuda) - ↑ Katherine L. Narr; Roger P. Woods; Paul M. Thompson; Philip Szeszko; Dilbert Robinson; Teodora Dimtcheva; Mala Gurbani; Arthur W. Toga; Robert M. Bilder (2007). «Relationships between IQ and Regional Cortical Grey Matter Thickness in Healthy Adults». Cerebral Cortex. 17 (9): 2163–2171. PMID 17118969. doi:10.1093/cercor/bhl125
- ↑ Alexandre F.M. DaSilva; Cristina Granziera; Josh Snyder; Nouchine Hadjikhani (2007). «Thickening in the somatosensory cortex of patients with migraine». Neurology. 69 (21): 1990–1995. PMC 3757544. PMID 18025393. doi:10.1212/01.wnl.0000291618.32247.2d
- ↑ Catharine Paddock (20 de novembro de 2007). «Migraine Sufferers Have Thicker Brain Cortex». Medical News Today. Arquivado do original em 11 de maio de 2008
- ↑ Datte R, Detre JA, et al. (outubro de 2011). «Absence of changes in cortical thickness in patients with migraine». Cephalalgia. 31 (14): 1452–8. PMC 3512201. PMID 21911412. doi:10.1177/0333102411421025
- ↑ Habib M (2000). «The neurological basis of developmental dyslexia: an overview and working hypothesis». Brain. 123 (12): 2373–99. PMID 11099442. doi:10.1093/brain/123.12.2373
- ↑ «Scientists identify a new kind of human brain cell». Allen Institute. 27 de agosto de 2018
- ↑ Shipp, Stewart (17 de junho de 2007). «Structure and function of the cerebral cortex». Current Biology. 17 (12): R443–9. PMC 1870400. PMID 17580069. doi:10.1016/j.cub.2007.03.044
- ↑ Meyer, Gundela; Goffinet, André M.; Fairén, Alfonso (1999). «Feature Article: What is a Cajal–Retzius cell? A Reassessment of a Classical Cell Type Based on Recent Observations in the Developing Neocortex». Cerebral Cortex. 9 (8): 765–775. PMID 10600995. doi:10.1093/cercor/9.8.765
- ↑ Judaš, Miloš; Pletikos, Mihovil (2010). «The discovery of the subpial granular layer in the human cerebral cortex». Translational Neuroscience. 1 (3): 255–260. doi:10.2478/v10134-010-0037-4
- ↑ Gilbert CD, Sigman M (2007). «Brain states: top-down influences in sensory processing». Neuron. 54 (5): 677–96. PMID 17553419. doi:10.1016/j.neuron.2007.05.019
|hdl-access=requer|hdl=(ajuda) - ↑ Cauller L (1995). «Layer I of primary sensory neocortex: where top-down converges upon bottom-up». Behav Brain Res. 71 (1–2): 163–70. PMID 8747184. doi:10.1016/0166-4328(95)00032-1
- ↑ Rubio-Garrido P, Pérez-de-Manzo F, Porrero C, Galazo MJ, Clascá F (2009). «Thalamic input to distal apical dendrites in neocortical layer 1 is massive and highly convergent». Cereb Cortex. 19 (10): 2380–95. PMID 19188274. doi:10.1093/cercor/bhn259
- ↑ Jones EG (1998). «Viewpoint: the core and matrix of thalamic organization». Neuroscience. 85 (2): 331–45. PMID 9622234. doi:10.1016/S0306-4522(97)00581-2
- ↑ Jones EG (1998). «Viewpoint: the core and matrix of thalamic organization». Neuroscience. 85 (2): 331–45. PMID 9622234. doi:10.1016/S0306-4522(97)00581-2
- ↑ Creutzfeldt, O. 1995. Cortex Cerebri. Springer-Verlag.
- ↑ a b Lam YW, Sherman SM (2010). «Functional Organization of the Somatosensory Cortical Layer 6 Feedback to the Thalamus». Cereb Cortex. 20 (1): 13–24. PMC 2792186. PMID 19447861. doi:10.1093/cercor/bhp077
- ↑ Pletikos, Mihovil; Sousa, Andre MM; et al. (22 de janeiro de 2014). «Temporal Specification and Bilaterality of Human Neocortical Topographic Gene Expression». Neuron. 81 (2): 321–332. PMC 3931000. PMID 24373884. doi:10.1016/j.neuron.2013.11.018
- ↑ Wolpert, Lewis (2015). Principles of development Fifth ed. UK: Oxford University Press. ISBN 9780199678143
- ↑ Warren N, Caric D, Pratt T, Clausen JA, Asavaritikrai P, Mason JO, Hill RE, Price DJ (1999). «The transcription factor, Pax6, is required for cell proliferation and differentiation in the developing cerebral cortex». Cerebral Cortex. 9 (6): 627–35. PMID 10498281. doi:10.1093/cercor/9.6.627
- ↑ Larsen, W J. Human Embryology 3rd edition 2001. pp 421-422 ISBN 0-443-06583-7
- ↑ Stephen C. Noctor; Alexander C. Flint; Tamily A. Weissman; Ryan S. Dammerman; Arnold R. Kriegstein (2001). «Neurons derived from radial glial cells establish radial units in neocortex». Nature. 409 (6821): 714–720. Bibcode:2001Natur.409..714N. PMID 11217860. doi:10.1038/35055553 Verifique o valor de
|name-list-format=amp(ajuda) - ↑ Sur, Mriganka; Leamey, Catherine A. (2001). «Development and Plasticity of Cortical Areas and Networks». Nature Reviews Neuroscience. 2 (4): 251–262. PMID 11283748. doi:10.1038/35067562
- ↑ a b Rakic, P (outubro de 2009). «Evolution of the neocortex: a perspective from developmental biology». Nature Reviews Neuroscience. 10 (10): 724–35. PMC 2913577. PMID 19763105. doi:10.1038/nrn2719
- ↑ Rakic, P (8 de julho de 1988). «Specification of cerebral cortical areas». Science. 241 (4862): 170–6. Bibcode:1988Sci...241..170R. PMID 3291116. doi:10.1126/science.3291116
- ↑ Fukuchi-Shimogori, T; Grove, EA (2 de novembro de 2001). «Neocortex patterning by the secreted signaling molecule FGF8». Science. 294 (5544): 1071–4. Bibcode:2001Sci...294.1071F. PMID 11567107. doi:10.1126/science.1064252
- ↑ Bishop, KM; Goudreau, G; O'Leary, DD (14 de abril de 2000). «Regulation of area identity in the mammalian neocortex by Emx2 and Pax6». Science. 288 (5464): 344–9. Bibcode:2000Sci...288..344B. PMID 10764649. doi:10.1126/science.288.5464.344
- ↑ Grove, EA; Fukuchi-Shimogori, T (2003). «Generating the cerebral cortical area map». Annual Review of Neuroscience. 26: 355–80. PMID 14527269. doi:10.1146/annurev.neuro.26.041002.131137
- ↑ Braitenberg, V and Schüz, A 1998. "Cortex: Statistics and Geometry of Neuronal Connectivity. Second thoroughly revised edition" New York: Springer-Verlag
- ↑ Saladin, Kenneth. Anatomy and Physiology: The Unity of Form and Function, 5th Ed. New York: McGraw-Hill Companies Inc., 2010. Print.
- ↑ Dorland's Medical Dictionary for Health Consumers, 2008.
- ↑ Yeo BT, Krienen FM, Sepulcre J, Sabuncu MR, Lashkari D, Hollinshead M, Roffman JL, Smoller JW, Zöllei L, Polimeni JR, Fischl B, Liu H, Buckner RL (2011). «The organization of the human cerebral cortex estimated by intrinsic functional connectivity». Journal of Neurophysiology. 106 (3): 1125–1165. PMC 3174820. PMID 21653723. doi:10.1152/jn.00338.2011
- ↑ Cathy J. Price (2000). «The anatomy of language: contributions from functional neuroimaging». Journal of Anatomy. 197 (3): 335–359. PMC 1468137. PMID 11117622. doi:10.1046/j.1469-7580.2000.19730335.x
- ↑ Kentar, Modar; Mann, Martina; Sahm, Felix; Olivares-Rivera, Arturo; Sanchez-Porras, Renan; Zerelles, Roland; Sakowitz, Oliver W.; Unterberg, Andreas W.; Santos, Edgar (15 de janeiro de 2020). «Detection of spreading depolarizations in a middle cerebral artery occlusion model in swine». Acta Neurochirurgica. 162 (3): 581–592. ISSN 0942-0940. PMID 31940093. doi:10.1007/s00701-019-04132-8
- ↑ Nakazawa T, Ohara T, Hirabayashi N, Furuta Y, Hata J, Shibata M, Honda T, Kitazono T, Nakao T, Ninomiya T (março de 2022). «Multiple-region grey matter atrophy as a predictor for the development of dementia in a community: the Hisayama Study». J Neurol Neurosurg Psychiatry. 93 (3): 263–271. PMC 8862082. PMID 34670843. doi:10.1136/jnnp-2021-326611
- ↑ Mukherjee, Raja A.S.; Hollins, Sheila (2006). «Fetal Alcohol Spectrum Disorder: An Overview». Journal of the Royal Society of Medicine. 99 (6): 298–302. PMC 1472723. PMID 16738372. doi:10.1177/014107680609900616
- ↑ Tarapore, PE; et al. (agosto de 2012). «Preoperative multimodal motor mapping: a comparison of magnetoencephalography imaging, navigated transcranial magnetic stimulation, and direct cortical stimulation.». Journal of Neurosurgery. 117 (2): 354–62. PMC 4060619. PMID 22702484. doi:10.3171/2012.5.JNS112124